Chlamydophila ( Chlamydia ) pneumonia. Медико-биологический Союз
Chlamydophila ( Chlamydia ) pneumonia
Манзенюк И. Н., Воробьева М. С., Государственный научно-исследовательский институт стандартизации и контроля медицинских иммунобиологических препаратов им. Л. А. Тарасевича.
Chlamydia pneumonia – патогенный облигатный внутриклеточный грамотрицательный микроорганизм, элементарные тельца которого при электронномикроскопическом исследовании имеют типичную грушеподобную форму, в отличие от округлых элементарных телец других видов хламидий. Штамм Chlamydia pneumonia получил название в результате слияния буквенных обозначений двух первичных изолятов TW-183 (изолирован из глаза ребенка в 1965 г. на Тайване) и AR-39 (идентифицирован из респираторного тракта в 1983 г. в США). Штаммы TWAR (Taiwan acute respiratory agent) выявлены только у человека, и изначально описывались как штаммы С.
Рис. 1. Цикл размножения хламидий.
Таксономия
Еще недавно С. pneumonia входила в порядок Chlamydiales, семейство Chlamydiaceae, род Chlamydia, который включал виды С. pneumonia, С. psittaci, С. trachomatis, С. percorum, и включала один серотип TWAR. Фундаментальные исследования в области систематики и генетики хламидий, проведенные Everett и др. [1, 2], полный сиквенс геномов С. trachomatis и С. pneumonia, анализ рибосомального оперона и открытие облигатных внутриклеточных микроорганизмов, имеющих схожий с хламидиями цикл развития, позволили уточнить таксономическое положение ряда представителей порядка Chlamydiales и предложить новую классификацию хламидий. Использование различных методов анализа – биохимического, серологических реакций, ДНК-анализа, сравнения протеиновых профилей, – а также различия в морфологии позволили по-новому разделить виды внутри рода хламидий.
ДНК-анализ последовательностей гена ompA указывает, что аминокислотный состав последовательностей основного белка наружной мембраны (МОМР) варьирует у всех штаммов С. pneumonia в пределах < 6 %, в то время как у других видов хламидий – >30 %.
Генетические (анализ ДНК-последовательностей генов рРНК, kdtA (KDO-трансфераза, 60 кДа-белка, богатого цистеином) и биологические различия позволили выделить биовары внутри вида С. pneumonia. Он объединяет три биовара – TWAR, Koala и Equine. Эталонным штаммом для С. pneumonia является TW-183T.
Умеренно патогенные изоляты биовара Koala выделены от сумчатых животных (Phascolarctos cinereus) в Австралии из глаз и мочевыводящих путей.
Биовар Equine вызывает асимптоматическую инфекцию и включает только один штамм N16, изолированный из респираторного тракта лошадей.
Широкий спектр заболеваний и синдромов, многие из которых имеют хроническое течение, связывают именно с С. pneumonia. Первоначально штаммы С. pneumonia были изолированы как респираторные патогены от людей. В дальнейшем были выделены штаммы С. pneumonia от коал и лошадей, по своим генетическим и антигенным характеристикам похожие на человеческие изоляты.
Заболеваемость данным патогеном во всем мире высокая, и большинство людей на протяжении жизни встречается с возбудителем неоднократно. С. pneumonia передается от человека к человеку воздушно-капельным путем. Отсутствуют случаи заражения от животных и птиц. Отмечено, что антитела к С. pneumonia у мужчин наблюдаются чаще, чем у женщин, в то время как у женщин преобладают антитела к С. trachomatis. Имеются некоторые сведения о периодичности инфекции: двух-трехлетний период увеличения числа заболевших инфекциями, вызванными С.
Молекулярно-биологические характеристики Chlamydia pneumonia
 pneumonia, выделенный от лошадей, содержит криптическую плазмиду (рСрпЕ1) размером 7.362 т. п. н. Данная плазмида полностью секвенирована, создана ее физическая и генетическая карты [б].
pneumonia, выделенный от лошадей, содержит криптическую плазмиду (рСрпЕ1) размером 7.362 т. п. н. Данная плазмида полностью секвенирована, создана ее физическая и генетическая карты [б]. Штаммы С. pneumonia содержит липополисахарид, МОМР, отр-2, отр-3, а также набор белков 90-100 кДа. Последние объединены в семейство протеинов, идентифицированных как Pomps или Pmps (полиморфные мембранные протеины). Большинство Pomps-генов было открыто при сиквенсе генома С. pneumonia и представляют собой гетерогенную группу из 21 гена с некоторыми общими характеристиками. В то же время протеиновый профиль С. pneumonia отличается от других представителей родов Chlamydia и Chlamydophila. Сравнительная оценка протеиновых полос различных штаммов С. pneumonia показала их идентичность, различия наблюдались лишь в количественных пропорциях. Все штаммы выявили наибольшее количество протеинов с молекулярным весом 98, 70, 54, 40 и 30 кДа. В иммуноблоте показано наличие у всех штаммов видоспецифическго протеина 54 кДа, играющего важную роль в иммунном ответе, а также показана реакция с 43 и 46 кДа протеинами [7].
С. pneumonia обладает системой секретирования III типа. Эта система является вирулентно ассоциированным механизмом, который не только осуществляет экспорт белков из грамотрицательных бактерий, но и усиливает их проникновение в цитозоль эукариотной клетки, в которой локализуется возбудитель, позволяя ему инвазировать хозяйскую клетку и разрушать ее защитные механизмы. Эта система известна для многих грамотрицательных патогенов растений и животных, таких как Yersinia, Shigella, Salmonella, энтеропатогенных Е. coli и Pseudomonas и т. д. Предполагают возможность секретирования данной системой в клетку хозяина фактора, ингибирующего апоптоз (запрограмированную гибель) этой клетки [10].
Клиника
Хламидии являются причиной разнообразной патологии у человека, которая включает не только хорошо известные и типичные для хламидийной инфекции трахому и урогенитальные заболевания (С. trachomatis), пневмонии (С. psittaci), инфекции верхнего респираторного тракта (С. pneumonia). Список заболеваний, которые, возможно, ассоциируются с хламидийной инфекцией, обширен и постоянно растет. Он включает острые и хронические заболевания типа астмы, рака легкого, артритов, саркоидоза и т. д. (таблица 2).
Таблица 2. Инфекционная патология у человека, вызываемая или ассоциируемая (в качестве кофактора) с Chlamydia pneumonia.
|
Патология респираторного тракта |
Патология сердечно-сосудистой системы |
Патология центральной нервной системы |
Патология других органов и тканей организма |
|
Синуситы Фарингиты Отиты Бронхиты Пневмонии Эмфизема Рак легкого Хроническая обструктивная легочная патология |
Острый инфаркт миокарда Реактивные васкулиты Аневризма Атеростклероз |
Болезнь Альцгеймера Множественный склероз Энцефалиты Менингоэнцефалиты Миелиты |
Реактивные артриты Саркоидоз Эритрема узловатая Острый лимфаденит Диабетическая нефропатия |
Недавно открытые хламидоподобные микроорганизмы (Simkaniae negevensis), образовавшие новое семейство в порядке Chlamydiales Simkaniaceae, вызывают пневмонии у взрослых и бронхиолиты у детей [12].
Патология респираторного тракта
Первичная связь С. pneumonia с разнообразной патологией верхних и нижних дыхательных путей у человека была установлена 10 лет назад. Данный возбудитель является важным этиологическим агентом респираторных заболеваний среди молодых людей, у которых инфекция часто протекает бессимптомно или в легкой форме и в ряде случаев остается нераспознанной. С. pneumonia является также причиной серьезных заболеваний дыхательного тракта у пожилых людей (пневмонии и бронхиты с выраженной симптоматикой). На долю этого вида хламидий приходится примерно 25 % всех случаев респираторных заболеваний, включая до 10 % эндемических пневмоний (до 50 % эпидемических), 5 % случаев острых и хронических бронхитов и 2 % фарингитов. Инкубационный период составляет приблизительно 21-30 дней [13, 14]. Возбудитель вызывает хронические риниты, фарингиты, острые отиты среднего уха [15], синуситы, гриппоподобные заболевания, обнаруживается в аденоидах у детей. В странах Европы и США инфекции, вызванные С. pneumonia, обычно не встречаются у детей в возрасте до 5 лет, но с высокой частотой роста (5-10 % ежегодно) выявляются у детей в возрасте 5-15 лет, а затем с более низкой частотой нарастания (1-2 % ежегодно) продолжает увеличиваться во всех возрастных группах. Антитела от первой инфекции сохраняются в течение 3-5 лет. К 20 годам более 50 % населения развитых стран имеют антитела к данному возбудителю. Прирост количества антител продолжается в течение всей жизни, достигая в пожилом возрасте до 80 % среди мужчин и до 70 % среди женщин. Возможно, это связано с хронизацией инфекции. Так, японские исследователи показали, что более часто инфекция встречается у подростков. Распространенность антител к С. pneumonia среди населения в Японии в среднем составляет 58.1 % [16]. Возможно, что в ряде случаев они свидетельствуют о наличии хронической инфекции [17, 18]. Титры антител к инфекциям, вызванным С. pneumonia, часто значительно превышают титры антител к инфекциям, обусловленным С.
В странах Европы и США инфекции, вызванные С. pneumonia, обычно не встречаются у детей в возрасте до 5 лет, но с высокой частотой роста (5-10 % ежегодно) выявляются у детей в возрасте 5-15 лет, а затем с более низкой частотой нарастания (1-2 % ежегодно) продолжает увеличиваться во всех возрастных группах. Антитела от первой инфекции сохраняются в течение 3-5 лет. К 20 годам более 50 % населения развитых стран имеют антитела к данному возбудителю. Прирост количества антител продолжается в течение всей жизни, достигая в пожилом возрасте до 80 % среди мужчин и до 70 % среди женщин. Возможно, это связано с хронизацией инфекции. Так, японские исследователи показали, что более часто инфекция встречается у подростков. Распространенность антител к С. pneumonia среди населения в Японии в среднем составляет 58.1 % [16]. Возможно, что в ряде случаев они свидетельствуют о наличии хронической инфекции [17, 18]. Титры антител к инфекциям, вызванным С. pneumonia, часто значительно превышают титры антител к инфекциям, обусловленным С. trachomatis. Антитела к C.trachomatis выявляются в сексуально активном возрасте и не превышают 20 % от общего количества населения. В менее развитых странах Азии и Африки TWAR-инфекция наблюдается и у детей в возрасте до 5 лет [13]. Предполагается, что каждый человек в течение жизни инфицируется этим микроорганизмом, многие даже неоднократно. Финскими исследователями показано, что инфекция, вызванная TWAR-штаммами, обуславливает менее тяжелое течение заболевания при последующей реинфекции [14]. Вопрос развития иммунитета к данному возбудителю спорный и требует дальнейшего изучения. Необъяснимым остается и факт более высокого выявления антител у мужчин старше 20 лет по сравнению с женщинами [14].
trachomatis. Антитела к C.trachomatis выявляются в сексуально активном возрасте и не превышают 20 % от общего количества населения. В менее развитых странах Азии и Африки TWAR-инфекция наблюдается и у детей в возрасте до 5 лет [13]. Предполагается, что каждый человек в течение жизни инфицируется этим микроорганизмом, многие даже неоднократно. Финскими исследователями показано, что инфекция, вызванная TWAR-штаммами, обуславливает менее тяжелое течение заболевания при последующей реинфекции [14]. Вопрос развития иммунитета к данному возбудителю спорный и требует дальнейшего изучения. Необъяснимым остается и факт более высокого выявления антител у мужчин старше 20 лет по сравнению с женщинами [14].
Важная особенность данного возбудителя – способность к персистенции. Кроме того, он способен вызывать патологию респираторного тракта в виде микст-инфекции – в ассоциации с другими респираторными патогенами [14]. С. pneumonia – потенциальный индуктор цитокинов; обладая биологически активным липополисахаридом, может индуцировать хроническую инфекцию в альвеолярных макрофагах, полученных методом in vitro. Причем включения, содержащие размножающийся патоген, способны сохраняться до 120 часов. In vitro на клеточных культурах С. pneumonia стимулирует продукцию бета-интерлейкина-1, гамма-интерферона и альфа-TNF, а также интерлейкина-8 и интерлейкина-6 [19]. Альвеолярные макрофаги отвечают на инфекцию окислительными реакциями, продукцией альфа-TNF, бета-интерпейкина-1 и интерлейкина-8. Установлено, что антимикробный ответ медиаторов не может уничтожить возбудителя и задержать его репликацию, но может усилить местный воспалительный ответ на С. pneumonia [20]. Попадая в кровь, возбудитель способен вызывать разнообразную экстрапульмональную патологию, становясь причиной кардитов, артритов и т. д.
Причем включения, содержащие размножающийся патоген, способны сохраняться до 120 часов. In vitro на клеточных культурах С. pneumonia стимулирует продукцию бета-интерлейкина-1, гамма-интерферона и альфа-TNF, а также интерлейкина-8 и интерлейкина-6 [19]. Альвеолярные макрофаги отвечают на инфекцию окислительными реакциями, продукцией альфа-TNF, бета-интерпейкина-1 и интерлейкина-8. Установлено, что антимикробный ответ медиаторов не может уничтожить возбудителя и задержать его репликацию, но может усилить местный воспалительный ответ на С. pneumonia [20]. Попадая в кровь, возбудитель способен вызывать разнообразную экстрапульмональную патологию, становясь причиной кардитов, артритов и т. д.
Рис. 2. Циклы развития Chlamidia.
С. pneumonia связывают с хронической ооструктивнои легочной патологией (ХОЛП), развитием эмфиземы легкого и астмы. Исследования с помощью ПЦР-анализа и выявления IgA (как маркера хронической инфекции) показали присутствие возбудителя в большинстве случаев хронической легочной инфекции.
Острая инфекция респираторного тракта, вызываемая С. pneumonia, может привести к развитию хронического астматического процесса из-за дальнейшей возможной хронизации инфекционного процесса и продуцирования в легких хламидийных компонентов, обладающих аллергенным действием [17]. Так, отмечено, что у взрослых в семь раз выше риск развития астмы после перенесенной инфекции, вызванной С. pneumonia. Ряд исследователей рекомендуют при диагностированной хронической астме провести обследование на предмет возможного хронического инфицирования С. pneumonia [21, 22]. В 18 контролируемых эпидемиологических исследованиях (более 4000 случаев) с использованием ПЦР-анализа, иммунофлюоресцентного метода для выявления антигена, ИФА для выявления специфических секреторных IgA (sIgA) и/или специфических сывороточных IgA, IgG и т. д. связь между инфицированием С. pneumonia и астмой обнаружена в 15 исследованиях. В восьми сообщениях и в описании еще 13 случаев инфекции С. pneumonia (всего более 100 больных) отмечено значительное улучшение, а в ряде случаев полное исчезновение симптомов астмы после пролонгированной антибиотикотерапии, направленной против С. pneumonia. Связь С. pneumonia с ХОЛП (более 1 000 случаев) была отмечена в пяти сообщениях из шести [13, 23]. У более чем 50 % детей, страдающих астмой, выявляются специфические IgE. Больные астмой имеют более высокие уровни IgA анти-Нsp60, чем больные с сердечно-сосудистой патологией. Полагают, что данные иммуноглобулины могут служить маркером для астматического процесса, вызванного С. pneumonia. Необходимы широкомасштабные эпидемиологические исследования для изучения роли этого патогена в патогенезе астмы.
pneumonia. Связь С. pneumonia с ХОЛП (более 1 000 случаев) была отмечена в пяти сообщениях из шести [13, 23]. У более чем 50 % детей, страдающих астмой, выявляются специфические IgE. Больные астмой имеют более высокие уровни IgA анти-Нsp60, чем больные с сердечно-сосудистой патологией. Полагают, что данные иммуноглобулины могут служить маркером для астматического процесса, вызванного С. pneumonia. Необходимы широкомасштабные эпидемиологические исследования для изучения роли этого патогена в патогенезе астмы.
С. pneumonia (TWAR), как и Mycoplasma pneumoniae, Legionella spp., C. psittaci, Coxiella burnetti и другие патогены, вызывает у людей атипичные пневмонии, характеризующиеся бронхо-легочным поражением, а также наличием других системных органных поражений. Для инфекции характерно клиническое течение средней тяжести, но возможно и тяжелое, с летальным исходом. Тяжелое течение чаще наблюдается у пожилых людей и у лиц с хроническими заболеваниями сердечнососудистой системы и респираторного тракта. Описаны спорадические случаи, эпидемические (особенно в закрытых коллективах) и внутрибольничные вспышки. Финскими исследователям описаны эпидемические вспышки среди рекрутов, продолжавшиеся пять-семь месяцев [24]. Соотношение заболевших – 60-80 на 1 000 мужчин. В доэпидемический период заболеваемость пневмонией, вызванной С. pneumonia, оценивалась как 1 на 1 000 человек. В среднем инкубационный период составил 28 дней (21 день для семейных пар). Среди студентов, больных пневмонией, С. pneumonia выявляют в 12 % случаев. Кроме того, С. pneumonia способна вызывать заболевания нижних дыхательных путей у ВИЧ-инфицированных больных [25].
Описаны спорадические случаи, эпидемические (особенно в закрытых коллективах) и внутрибольничные вспышки. Финскими исследователям описаны эпидемические вспышки среди рекрутов, продолжавшиеся пять-семь месяцев [24]. Соотношение заболевших – 60-80 на 1 000 мужчин. В доэпидемический период заболеваемость пневмонией, вызванной С. pneumonia, оценивалась как 1 на 1 000 человек. В среднем инкубационный период составил 28 дней (21 день для семейных пар). Среди студентов, больных пневмонией, С. pneumonia выявляют в 12 % случаев. Кроме того, С. pneumonia способна вызывать заболевания нижних дыхательных путей у ВИЧ-инфицированных больных [25].
Не исключено, что С. pneumonia отягощает патогенез хронического бронхита. Следует отметить, что в ряде случаев интенсивный курс лечения, направленный на удаление возбудителя, стабилизирует течение заболевания. При хроническом бронхите выявление IgG не всегда является достоверным маркером, т. к. инфекция диагностируется менее чем в 10% случаев. Выявление специфических IgA в сыворотке, наличие специфических секреторных IgA и ДНК в слюне свидетельствует о возможности присутствия данного возбудителя в легких более чем у 50% больных хроническим бронхитом, особенно у курильщиков, снижение клеточного иммунитета которых способствует инфицированию С. pneumonia [17].
Выявление специфических IgA в сыворотке, наличие специфических секреторных IgA и ДНК в слюне свидетельствует о возможности присутствия данного возбудителя в легких более чем у 50% больных хроническим бронхитом, особенно у курильщиков, снижение клеточного иммунитета которых способствует инфицированию С. pneumonia [17].
Ряд исследователей отмечают, что инфекция, вызываемая С. pneumonia, достаточно часто выявляется у курящих больных раком легкого. Считают, что данный возбудитель может быть независимым фактором риска в развитии рака легкого. У курящих больных раком С. pneumonia диагностируется в 90% случаев. В результате иммуносупрессионного эффекта курения в ряде случаев на фоне хронического бронхита активизируется хроническая инфекция, вызываемая С. pneumonia. Специфические IgA обнаруживались значительно чаще в группе больных раком легкого, чем в контрольных группах, включающих группу здоровых и группу больных с сердечно-сосудистой патологией, а также лиц с таким фактором риска, как курение [26, 27].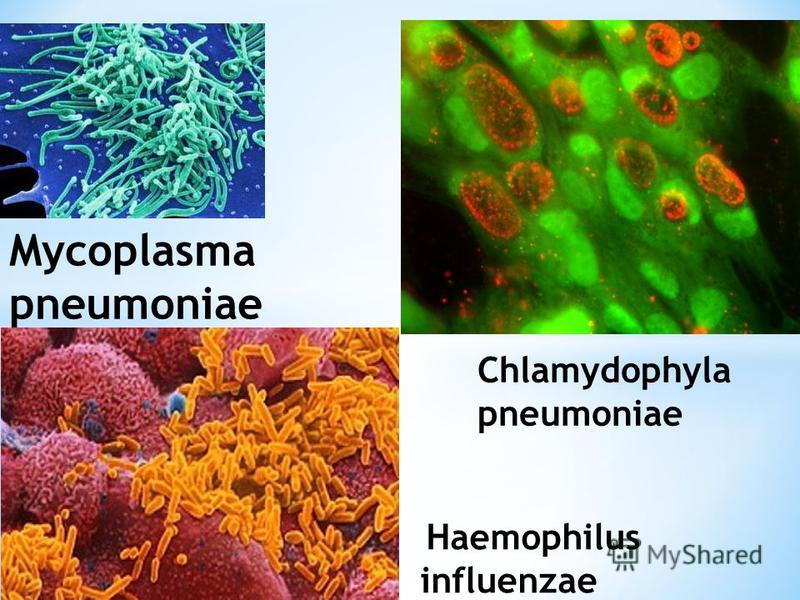
Сердечно-сосудистая патология
Многочисленными исследованиями установлена связь хронической инфекции С. pneumonia как фактора риска с развитием сердечной патологии. Это подтверждается сероэпидемиологическими исследованиями, наличием белков данного возбудителя в циркулирующих иммунных комплексах и присутствием ЛПС-содержащих иммунных комплексов. Хроническая хламидийная инфекция кардиоваскулярной системы может быть связана с атеросклерозом, аневризмой аорты, острым инфарктом миокарда, с хронической сердечно-сосудистой патологией, миокардитами [28-33]. Японскими исследователями показана взаимосвязь между развитием гипертензии у людей и наличием инфекции, вызванной С. pneumonia [34]. Установлена высокая частота персистирования С. pneumonia в верхних отделах респираторного тракта у пациентов с ишемической болезнью сердца [35]. В то же время неясно, служит ли толчком к развитию болезни персистирующая инфекция или реинфекция [36]. Известно, что инфекция может воздействовать на липидный обмен. В отличие от пневмоний, вызванных другими возбудителями, при острой пневмонии, вызванной С pneumonia, отмечено снижение количества высоконасыщенных липопротеинов и увеличение триглицеридов [37]. Попытка связать наличие IgG к С. pneumonia и высокого уровня липопротеинов в крови с острыми мозговыми нарушениями не привела к успеху [38]. В отдельных работах показана возможная связь между атерогенным липидным профилем и наличием специфических антител к С. pneumonia. Возможно, изменения в липидном профиле могут лежать в основе связи С. pneumonia с ишемической болезнью сердца [39], что было продемонстрировано в случаях ангиографически подтвержденной ишемической болезни сердца [40]. Как показали исследования, сердечно-сосудистая патология, опосредованная С. pneumonia, индуцируется антигенной мимикрией возбудителя к специфическому протеину сердечной мышцы. Установлена гомология нуклеотидных последовательно-стей тяжелой цепи сердечного специфичного мышечного альфа-миозина с богатым цистеином мембранным протеином 60 кДа С.
В отличие от пневмоний, вызванных другими возбудителями, при острой пневмонии, вызванной С pneumonia, отмечено снижение количества высоконасыщенных липопротеинов и увеличение триглицеридов [37]. Попытка связать наличие IgG к С. pneumonia и высокого уровня липопротеинов в крови с острыми мозговыми нарушениями не привела к успеху [38]. В отдельных работах показана возможная связь между атерогенным липидным профилем и наличием специфических антител к С. pneumonia. Возможно, изменения в липидном профиле могут лежать в основе связи С. pneumonia с ишемической болезнью сердца [39], что было продемонстрировано в случаях ангиографически подтвержденной ишемической болезни сердца [40]. Как показали исследования, сердечно-сосудистая патология, опосредованная С. pneumonia, индуцируется антигенной мимикрией возбудителя к специфическому протеину сердечной мышцы. Установлена гомология нуклеотидных последовательно-стей тяжелой цепи сердечного специфичного мышечного альфа-миозина с богатым цистеином мембранным протеином 60 кДа С. pneumonia, что может индуцировать аутоиммунные процессы в сердечной патологии [41]. Установлено, что инфекция, вызываемая С. pneumonia, ведет к репрессии антикоагулянтного протеина – тромбомодулина на поверхности эндотелиальных клеток, выстилающих стенки сосудов. Нарушение гомеостаза эндотелия в ответ на инфекцию по отношению к прокоагулянтам может приводить к увеличению локального тромбообразования и коронарной болезни сердца [42].
pneumonia, что может индуцировать аутоиммунные процессы в сердечной патологии [41]. Установлено, что инфекция, вызываемая С. pneumonia, ведет к репрессии антикоагулянтного протеина – тромбомодулина на поверхности эндотелиальных клеток, выстилающих стенки сосудов. Нарушение гомеостаза эндотелия в ответ на инфекцию по отношению к прокоагулянтам может приводить к увеличению локального тромбообразования и коронарной болезни сердца [42].
Сероэпидемиологические исследования, проведенные в ряде индустриально развитых стран мира и основанные на исследовании специфических IgG и IgA, специфических иммунных комплексов, показали, что острый инфаркт миокарда у абсолютного большинства больных связан с сероконверсией против ЛПС-эпитопов С. pneumonia [43], что может быть следствием обострения хронической болезни, вызванной С. pneumonia [17] IgG1 является наиболее общим подклассом среди антител к ЛПС С. pneumonia [44]. У пациентов с острым инфарктом миокарда обнаруживаются более высокие показатели значений специфических IgA./mycoplasma-bacteria--illustration-758305183-5ad4fdabae9ab80038b98054.jpg) Вместе с тем, необходимы более углубленные исследования, чтобы оценить роль внутриклеточных патогенов, прежде всего С. pneumonia, в развитии острых сердечнососудистых синдромов [45].
Вместе с тем, необходимы более углубленные исследования, чтобы оценить роль внутриклеточных патогенов, прежде всего С. pneumonia, в развитии острых сердечнососудистых синдромов [45].
Рис 3. Электронная микроскопия. Chlamidia pneumonia в коронарной артерии. Внутри пенистых клеток в ткани взятой ихз атеромы коронарной артерии эндосомы содержат грушевидные элементарные тельца C. Pneumonia (Cambell L.A. et al., 2001).
Из представленных выше данных следует, что необходимо проведение широкомасштабных клинических исследований, направленных на оценку эффективности применения антихламидийных антибиотиков в лечении ишемической сердечно-сосудистой патологии, с учетом возможной роли Chlamydia pneumonia в атерогенезе. Определенная связь этих факторов подтверждается многочисленными эпидемиологическими и гистологическими исследованиями, в том числе и на животных [46-51].
Три небольших пилотных испытания по изучению эффективности применения этиотропной терапии в лечении ишемической сердечно-сосудистой патологии уже были проведены. В двух из них была подтверждена положительная роль антибиотикотерапии в улучшении состояния пациентов с сердечно-сосудистой патологией, в одном исследовании положительный эффект не отмечен. Из-за небольшого числа пациентов, взятых в эти испытания, нельзя достоверно ответить, является ли применение антибиотикотерапии у больных с сердечно-сосудистой патологией удачной. В настоящее время продолжаются по крайней мере два подобных клинических испытания, которые могут дать ответ на вопрос: “может ли применение антибиотиков замедлить развитие коронарной патологии”. Это WIZARD-испыта-ния, спонсируемые “Pfizer Co”, и ACES-испытания, проводимые “National Heart Lung and Blood Institute”. В испытаниях участвуют 3500 – 4000 пациентов с доказанной сердечно-сосудистой патологией, которые наблюдаются на протяжении четырех лет с момента начала испытаний. В качестве антибиотика в обоих испытаниях используется азитромицин. В WIZARD-испытаниях пациенты получают 600 мг азитромицина и плацебо однократно в неделю в течение 3 месяцев.
В двух из них была подтверждена положительная роль антибиотикотерапии в улучшении состояния пациентов с сердечно-сосудистой патологией, в одном исследовании положительный эффект не отмечен. Из-за небольшого числа пациентов, взятых в эти испытания, нельзя достоверно ответить, является ли применение антибиотикотерапии у больных с сердечно-сосудистой патологией удачной. В настоящее время продолжаются по крайней мере два подобных клинических испытания, которые могут дать ответ на вопрос: “может ли применение антибиотиков замедлить развитие коронарной патологии”. Это WIZARD-испыта-ния, спонсируемые “Pfizer Co”, и ACES-испытания, проводимые “National Heart Lung and Blood Institute”. В испытаниях участвуют 3500 – 4000 пациентов с доказанной сердечно-сосудистой патологией, которые наблюдаются на протяжении четырех лет с момента начала испытаний. В качестве антибиотика в обоих испытаниях используется азитромицин. В WIZARD-испытаниях пациенты получают 600 мг азитромицина и плацебо однократно в неделю в течение 3 месяцев. В ACES-испытаниях лечение продолжается в течение года. Столь длительное лечение обусловлено трудностями в успешном лечении хронической хламидийной инфекции. Судить об эффективности лечения будут по количеству летальных случаев, обусловленных сердечнососудистой патологией, инфарктов миокарда в опытной и контрольной группах, показателях реконвалесценции, госпитализации и т. д. В то же время до получения окончательных результатов этих испытаний не рекомендуется использовать антибиотикотерапию для предотвращения осложнений сердечно-сосудистой патологии и атеросклероза.
В ACES-испытаниях лечение продолжается в течение года. Столь длительное лечение обусловлено трудностями в успешном лечении хронической хламидийной инфекции. Судить об эффективности лечения будут по количеству летальных случаев, обусловленных сердечнососудистой патологией, инфарктов миокарда в опытной и контрольной группах, показателях реконвалесценции, госпитализации и т. д. В то же время до получения окончательных результатов этих испытаний не рекомендуется использовать антибиотикотерапию для предотвращения осложнений сердечно-сосудистой патологии и атеросклероза.
Атеросклероз сердечно-сосудистой системы является основной причиной смертности населения в развитых странах. Выявлен ряд факторов риска атеросклеротического поражения сосудов, к числу которых относятся наследственность, гиперхолестеринемия, нарушенный обмен веществ, артериальная гипертензия, курение, сахарный диабет, ожирение и гиподинамия. Кроме того, есть определенные основания предполагать, что одним из таких факторов может являться инфекционный процесс. Бактериальная теория атеросклероза, впервые выдвинутая еще более ста лет назад, вновь приобретает все больший научный и практический интерес и имеет ряд доказательств.
Рис.4. Иммуногисто-химическое окрашивание. С.pneumonia в фибролипидных бляшках из атеромы коронарной артерии. А – положительное окрашивание специфическими моноклональными антителами. Пенистые клетки (из фибролипидных бляшек), содержащие С.pneumonia. В – отрицательное окрашивание нормального контрольного образца (Cambell LA. et al,, 2001).
По данным ряда авторов [28, 29, 33, 52, 53], атеросклероз может быть результатом хронической персистентной инфекции, вызванной С. pneumonia. Главными аргументами в пользу хламидийной этиологии этого заболевания являются корреляция сердечно-сосудистой патологии и других атеросклеротических поражений с динамикой выявления специфических антител против С. pneumonia, что было показано в сероэпидемических исследованиях, проведенных в Финляндии, Швеции, США, Германии, Великобритании и Нидерландах и основанных на измерении IgG и IgA или специфических иммунных комплексов [54]. К настоящему времени проведено около 50 сероэпидемиологических исследований по выявлению TWAR-антител при различных атеросклеротических заболеваниях. Более чем в 40 исследованиях обнаружена связь между присутствием специфических антител и заболеванием, в некоторых исследованиях она не прослеживалась. Частота выявления антител к С. pneumonia у лиц с доказанной сердечно-сосудистой патологией составляла в проведенных исследованиях 80-85 %. При этом в ряде случаев отмечено отсутствие специфических антител у пожилых людей с атеросклерозом [55].
Рис. 5. Течение инфекции в атеросклерозе перед реактивным артритом.
Данный возбудитель удается выявить различными методиками (ПЦР, иммуногистология, электронная микроскопия, культуральный метод) из тканей атеросклеротических бляшек стенок артерий, при отсутствии такового в нормальной ткани стенок артериальных артерий [13, 17, 18, 56, 57]. Показана возможность инфицирования и размножения С. pneumonia в клетках-мишенях атеросклероза (эндотелий, макрофаги, мышечные клетки) в опытах in vitro при исследовании эндотелиальных клеток сосудов человека, а также при моделировании инфекции у животных [19, 58]. Отмечено, что хламидийный белок теплового шока (Hsp 60) обнаруживается в атеросклеротической бляшке в значительном количестве вместе с аналогичным эндогенным белком человека. Этот белок может являться активным важную роль в атерогенезе, стимулируя выработку макрофагами альфа-фактора некроза опухоли и экспрессию металлопротеиназ, желатиназы 92 кДа и способствуя разрушению тканей сосудов [59, 60]. Показано, что эндотелиальные клетки в ответ на проникновение С. pneumonia увеличивают экспрессию генов цитокинов (интерлейкин-1), хемокинов (интерлейкин-8 и моноцитарный хемотаксический протеин 1), клеточных факторов роста (гепаринсвязанный эпидермальноподобный фактор, основной фибробластный фактор и т.д.), внутриклеточных киназ и поверхностных клеточных рецепторов с сигнальной транедуцирующей активностью [61]. Цитокиновый ответ на проникновение С. pneumonia, по-видимому, может играть роль в развитии локального воспаления в местах инфекции (сосуды) [19]. А ЛПС С. pneumonia стимулирует накопление макрофагами эфиров холестерина в присутствии липопротеидов низкой плотности, индуцируя образование “пенистых” клеток. Экспериментально доказана способность штаммов С. pneumonia индуцировать атеросклеротические изменения в клетках стенок сосудов у кролика [62-64].
С. pneumonia способна инфицировать несколько типов клеток, включая моноцитпроизводные макрофагов, играющих существенную роль в развитии атеросклероза. Однако персистенция С. pneumonia в мононуклеарных клетках плохо изучена [65]. Имеется ряд сообщений, подтверждающих присутствие ДНК С. pneumonia в периферийных мононуклеарных клетках крови у пациентов с коронарной ангиографией, причем чаще у курильщиков [66]. С. pneumonia может участвовать в локальном иммунном ответе и через инфицированные макрофаги расширять зоны атеросклеротических повреждений. Хроническая инфекция С. pneumonia у нормолипидных мышей индуцирует воспалительные изменения, но не атеросклеротические изменения стенок сосудов. На этой же лабораторной модели инфекции показано, что атерогенный эффект С. pneumonia развивается при наличии гиперлипидемии [67]. Показано, что на фоне диетиндуцируемой гиперлипидемии у мышей с дефицитом LDL-рецепторов происходит индукция и развитие атерогенеза, которое может быть обусловлено инфекцией, вызываемой С. pneumonia [68]. В то же время лечение азитромицином не уменьшает размер развившихся атеросклеротических повреждений в экспериментах на инфицированных мышах. Предполагают, что лечение острой инфекции С. pneumonia антихламидийными препаратами, возможно, оказывает профилактическое противоатерогенное действие данного возбудителя [69].
Рис. 6. Возможные механизмы вовлечения C. pneumonia в атерогенез.
Таким образом, исследования различных авторов показывают, что колонизация С. pneumonia стенок артерии при хроническом инфекционном процессе приводит к дисфункции и нарушению гомеостаза эндотелия сосудов, способствуя развитию атеросклеротического процесса и хронической сердечно-сосудистой патологии. Этому, возможно, способствует неоднократное в течение жизни реинфицирование человека С. pneumonia, которое приводит к активации вялотекущего аутоиммунного процесса, Т-клеточного звена иммунитета, поддерживая таким образом интрамуральный хронический воспалительный процесс.
Возможно, что не только С. pneumonia, но и другие возбудители в атеросклеротической бляшке, таких как цитомегаловирус, вирус герпеса, Porphyromonas gingivalis и Streptococcus sanguis, могут вызывать повреждение клеток сосудов, приводя в дальнейшем, в ассоциации с классическими факторами риска, к развитию сердечно-сосудистой патологии и появлению атероскперотических изменений в сосудах [70-72].
Известно, что аневризма брюшной аорты чаще диагностируется у пожилых курящих мужчин с хроническим бронхитом. С помощью иммуногистохимического анализа удалось детектировать ЛПС хламидий в аневризме брюшной аорты в биоптатном материале у пациентов после операции. У большинства пациентов удалось детектировать белковые антигены С. pneumonia и ДНК возбудителя с помощью иммунологических методов исследования и ПЦР-анализа [73| Отмечено частое присутствие С. pneumonia в измененных тканях брюшной аорты при аневризме в живой, репликативной форме. Данные больные имеют более высокие серологические маркеры для инфекции С. pneumonia, чем в контроле. Аневризма стенки сосуда ха рактеризуется дегенерацией эластина и коллагена, а также инфильтрацией ее макрофагами и В-клетками [74]. У изолятов С. pneumonia установлена протеазная активность, которая способствует деструктивному действию данного возбудителя на стенки сосудов.
Патология центральной нервной системы
Рядом авторов показано развитие неврологических осложнений у больных с инфекциями, вызванными С. trachomatis и С psittaci. Имеется ряд сообщений о возможной связи нервной патологии с инфицированном хламидиями, в частности сообщается о роли С. pneumonia в возникновении острых инфекций ЦНС: энцефалиты, менингиты, менингоэнцефалиты, миелиты и синдрома Guillain-Barre.
Предполагают, что С. pneumonia играет роль в развитии цереброваскулярной патологии, особенно если наблюдается связь с курением, скоротечными ишемическими приступами, а также на фоне развития атеросклероза каротидной артерии. Отмечено четырехкратное увеличение риска развития цереброваскулярной болезни при наличии инфекции С. pneumonia [76].
В последние годы появляются сообщения о роли С. pneumonia в развитии наиболее важных патологий в ЦНС – множественного склероза и болезни Альцгеймера. Имеется ряд непрямых доказательств о возможной инфекционной и воспалительной этиологии болезни Альцгеймера. При сравнительном изучении патологоанатомических образцов головного мозга от пациентов в последней стадии болезни Альцгеймера и от контрольной группы (отсутствие данного заболевания) американскими исследователями установлена возможная связь С. pneumonia с болезнью Альцгеймера. При использовании широкого арсенала современных диагностических методов (ПЦР-анализ, включая RT-ПЦР; электронную и иммуноэлектронную микроскопию, иммуногистохимию и иммунозонды, а также культуральные методы исследования) было показано наличие живого (элементарные, ретикулярные тельца и ДНК хламидий, положительные культуральные высевы) и транскрипционно активного (транскрипты РНК двух важнейших генов) возбудителя С. pneumonia в участках головного мозга, пораженного болезнью Альцгеймера [77, 78]. Ряд других исследований подтверждают связь С. pneumonia с tau-ассоциированной нейрофибриллярной патологией [79]. В других исследованиях показано, что мозг, пораженный болезнью Альцгеймера, содержит незначительное количество копий ДНК С. pneumonia. По-видимому, различия, обнаруживаемые в исследованиях различных авторов, могут быть обусловлены низкой чувствительностью существующих методов ПЦР-анализа, недостаточным количеством образцов, взятых из различных мест мозга, пораженного данной патологией. Этими же исследователями отмечено, что в 85.7% (18 из 21) случаях с помощью ПЦР-анализа подтверждено обнаружение ДНК С. pneumonia в постмортальных образцах мозга, пораженных болезнью Альцгеймера, при отрицательных данных в контрольной группе (0 из 10). Авторы указывают на необходимость повышения чувствительности существующих методов ПЦР-анализа для обнаружения ДНК С. pneumonia. Данные исследования предполагают, что С. pneumonia может быть кофактором развития последней стадии болезни Альцгеймера.
Множественный склероз – одно из наиболее распространенных демиелинизирующих заболеваний человека. Известно, что многие факторы могут быть вовлечены в патогенез развития множественного склероза. Этиология данного заболевания до настоящего времени окончательно не определена. Имеются данные, аргументирующие возможную инфекционную этиология множественного склероза. В своей работе Sriam et al. [80] сообщает о возможной связи С. pneumonia с множественным склерозом. Приведены данные о первом случае связанного с С. pneumonia множественного склероза, при котором наблюдаются ли клиническое улучшение на фоне терапии антихламидийными препаратами. В другом исследовании с помощью ПЦР-анализа, серологических и культуральных методов исследования показано преобладание С. pneumonia в спинномозговой жидкости у больных с множественным склерозом по сравнению с контрольными группами. В то же время имеются данные других исследователей, показавшие отрицательные результаты в ПЦР-анализе и культуре клеток при изучении образцов спинномозговой жидкости от 99 больных (48 с множественным склерозом и 51 контроль) или низкие показатели обнаружения ДНК С.pneumonia с помощью ПЦР-анализа при изучении образцов спинномозговой жидкости от 37 больных множественным склерозом (только в 13%) и отсутствие ее в контрольной группе [81, 82]. При серологическом исследовании другими авторами показано наличие антител к С. pneumonia в спинномозговой жидкости у 8 из 22 больных множественным склерозом. Сделана попытка изоляции культуры С. pneumonia из образцов спинномозговой жидкости у больных с множественным склерозом, что удалось только в 1.7 % случаев [83]. Анализ проведенных исследований в целом может свидетельствовать о возможности того, что инфекция С. pneumonia является пусковым механизмом аутоиммунных процессов у больных множественным склерозом, а выявление возбудителя различными методами может зависеть от стадии заболевания. Необходимо отметить, что определение роли С. pneumonia в развитии множественного склероза требует дополнительных исследований.
Другая патология
Хламидии могут размножаться в клетках ретикулоэндотелиальной системы. В связи с этим некоторые исследователи предполагают наличие возможной связи между хламидиями и лимфомой. Так, известно, что Helicobacter pylori связана с В-клеточной лимфомой желудка. В одной исследовательской работе показано, что у пациентов с лимфомой значительно чаще выявляются специфические иммунные комплексы, связанные с С. pneumonia и С. trachomatis, чем в контрольной группе [84]. В то же время отсутствуют четкие данные, подтверждающие связь между хламидийной инфекцией и развитием лимфомы. Некоторые из представителей хламидий, например биовар венерической лимфогранулемы С. trachomatis, может явиться причиной гигантских клеточных преобразований и хронического гранулематоза.
Имеется ряд сообщений, посвященных связи С. pneumonia с саркоидозом и хроническим гранулематозным воспалением, реактивными васкулитами, цистическим фиброзом, с узловатой эритемой, реактивными артритами [13], острым лимфаденитом [85]. Японскими исследователями обнаружена взаимосвязь хронической инфекции С. pneumonia с диабетической нефропатией. Ими же показано более чем четырехкратное увеличение риска развития нефропатии в группах больных, имеющих высокий процент выявляемости антител к С. pneumonia [86]. В таблице 2 эти данные обобщены. Типично, что все эти нарушения, как правило, возникают после инфицирования респираторного тракта С. pneumonia [17].
Ряд исследований подтверждает, что хламидии в суставах присутствуют в измененной, персистирующей и труднокультивируемой форме. С. pneumonia может играть роль в патогенезе некоторых артритов [87]. При поражении суставов ДНК С. pneumonia обнаруживают в синовиальной жидкости приблизительно в 20% исследований. Однако необходимо отметить, что данный микроорганизм играет значительно меньшую роль при артритах, чем С. trachomatis [88].
Диагностика
Диагностика как острой, так и хронической (персистирующей) инфекции С. pneumonia затруднена из-за особенностей ее течения, низкой концентрации возбудителя в месте инфекции, наличием перекрестных серологических реакций. К сожалению, на сегодняшний день отсутствуют стандартизованные методы в выявлении антигенов, антител и ДНК С. pneumonia отмечены значительные различия в результатах исследовании при использовании тест-систем различных производителей. В связи с этим актуальной в настоящее время является проблема стандартизации технологий производства, методов применения и качества препаратов, используемых в постановке диагноза инфекции, вызванной С. pneumonia.
Культуральный метод диагностики, достаточно хорошо зарекомендовавший себя при диагностике инфекций, вызываемых С. trachomatis, менее эффективен при изоляции С. pneumonia. Отмечено, что возбудитель плохо размножается в культуре клеток, обычно используемых для культивирования хламидий, а некоторые штаммы С. pneumonia не размножаются в куриных эмбрионах, в связи с чем многие лаборатории мира имеют большие трудности как при изоляции возбудителя из клинических образцов при острой инфекции, так и при дальнейшем культивировании С. pneumonia. При хронических формах инфекции С. pneumonia в большинстве случаев культуральный метод диагностики практически дает отрицательные результаты. В настоящее время не выявлена культура клеток, наиболее чувствительная для изоляции штаммов С. pneumonia. Чаще всего для этой цели используются перевиваемые линии клеток HL и Нер-2. Имеются лишь отдельные сообщения об успешной изоляции штаммов С. pneumonia от пациентов с бессимптомной формой инфекции и негативной серологией. Показано, что успешной изоляции возбудителя можно достигнуть при позитивной сероконверсии в 50% случаев. Имеются данные, что штаммы С. pneumonia у различных групп населения могут обладать различным клеточным тропизмом и активностью размножения в культуре клеток. Необходима стандартизация культурального метод диагностики, особенно в отношении наиболее чувствительны линий клеток, условий инфицирования, пассирования и идентификации выделенных штаммов хламидий [89].
Иммуноферментный анализ используют для выявления общего группового ЛПС-антигена, который присутствует у всех хламидий. Перспективным является метод иммуногистохимического окрашивания биопсийных препаратов тканей с использованием специфических моноклональных антител к С. pneumonia и стрептавидин-биотиновой технологии [90].
Иммунофлюоресцентный анализ, направленный на выявление антигенов С. pneumonia, не обладает высокой чувствительностью при анализе фарингеальных мазков. Из-за разнообразия фарингеальной микрофлоры поиск элементарных телец С. pneumonia в образцах затруднен. Чувствительность данного метода составляет 20-60%, а специфичность зависит от навыков персонала.
Серологические методы исследования наиболее широко применяются в диагностике инфекций, вызываемых С. pneumonia. Первую связь С. pneumonia с болезнью у человека продемонстрировал Saikku [14], используя микроиммунофлюоресцентный метод (МИФ), которой является золотым стандартом в серологии. В то же время имеется ряд проблем, связанных с использованием МИФ-серологии: нестандартность реагентов, методов оценки результатов анализа, различия используемых диагностических критериев. Описаны значительные межлабораторные вариации в результатах МИФ-диагностики, в частности при оценке титров исследуемых сывороток. Наиболее результативной является комплексная диагностика при использовании различных методов. Показано, что четырехкратное увеличение титров антител к С. pneumonia в комбинации с результатами ПЦР-анализа или культурального метода – наилучший диагностический критерий наличия острой инфекции. При серодиагностике присутствие IgM-антител подтверждает острую или недавнюю инфекцию, высокие титры к IgA и/или IgG подтверждают наличие острой или манифестной хронической инфекции. Антитела класса М вырабатываются, как правило, на ЛПС С. pneumonia при первичной инфекции, а антитела класса А являются маркерами реинфицирования. Детекция специфических IgG в одном образце сыворотки имеет ограниченную диагностическую ценность. Только значительное увеличение титров специфических антител класса G в парных сыворотках дает возможность предполагать наличие острой или манифестной инфекции С. pneumonia. Серологическое исследование, проводимое без ДНК-анализа или культурального метода диагностики, а также при отсутствии парных сывороток, носит ретроспективный анализ.
Классический метод диагностики – реакция связывании комплемента – может быть использована в серологической ни агностике первоначальной инфекции, однако этот тест малочувствителен у пациентов с реинфекциями, может давать перекрестные реакции с другими видами хламидий, такими как С. psittaci.
В последние годы разработаны специфические ELISA тест-системы, направленные на выявление видоспецифических антител классов G, А, М к С. pneumonia. Эти тест-системы в настоящее время повсеместно используются в клинической практике. При диагностике с применением серологических методов исследователь сталкивается с трудностями интерпретации получаемых результатов. Так как антитела к С. pneumonia обычно присутствуют в старшей возрастной группе и у здоровых, то соотнесение их с определенной стадией болезни может быть затруднено. В то же время продолжительное присутствии короткоживущих IgA может служить индикатором манифестации хронической инфекции. Отмечено, что титры IgG к С. pneumonia были выше в сыворотках с сопутствующими IgA-антителами. IgA никогда не детектировались в сыворотке без специфических IgG. IgG1 являются наиболее распространенным субклассом G-антител к С. pneumonia [91]. При инфекциях, осложненных С. trachomatis, титры антител к С. pneumonia часто меньше, чем титры антител к С. trachomatis. Наличие высоких титров IgG к С. pneumonia в контрольных группах, возможно, связано с высоким уровнем общего IgG. Наилучший маркер хронической инфекции С. pneumonia – иммунные комплексы и IqA [92].
ПЦР-анализ, как и другие методы амплификации нуклеиновых кислот, является потенциально ценным диагностическим инструментом, который способен улучшить диагностику не только ранней, но и хронической (персистирующей) инфекции, вызываемой С. pneumonia. Клиническими образцами для ДНК анализа могут служить назофарингеальные смывы, ткани пораженных сосудов или органов, мононуклеарные клетки периферической крови. При диагностике с помощью ПЦР-анализа следует учитывать интервал между развитием клинических симптомов и забором образца, при этом чем больше интервал, тем вероятнее обнаружение ДНК в мононуклеарных клетках. Это является маркером возможного присутствия С. pneumonia в сосудистой стенке, в частности в атеросклеротической бляшке. Кроме того, этот метод может быть новым “золотым стандартом” для диагностики хронической инфекции С. pneumonia у больных ХОЛП – как маркер присутствия возбудителя в нижних отделах респираторного тракта. Однако необходимо отметить, что ДНК-методы амплификации С. pneumonia недостаточно стандартизованы. Необходима отработка и оптимизация процедур выделения нуклеиновых кислот из различных клинических образцов (возможно, нескольких), проведение различных методов ДНК-амплификаций [93]. Отмечены существенные различия в оценке чувствительности и специфичности ПЦР-анализа, используемого для выявления ДНК С. pneumonia в клинических образцах [94]. Необходима отработка методов экстракции ДНК С. pneumonia из ткани сосудов, удаление соответствующих ингибиторов ПЦР-реакции. Большое значение в ДНК-диагностике инфекций, вызванных С. pneumonia, приобретают методы ДНК-амплификаций, которые легко автоматизировать и стандартизовать, в частности, SDA-метод [95] и новый метод DIG-PCR-EIA, который имеет большую чувствительность по сравнению с обычной двухпраймерной или nested-ПЦР [96].
Чувствительность к антибактериальным препаратам. Лечение.
Открытие новых возбудителей требует разработки новых методов в диагностике и лечении малоизученных хронических заболеваний. С. pneumonia не является исключением. Обобщенные сведения о чувствительности штаммов С. pneumonia к антибактериальным препаратам представлены в таблице 3 [97].
Таблица 3. Чувствительность штаммов С. pneumonia к антибактериальным препаратам.
|
Антибактериальные препараты |
МПК90, мкг/мл |
|
Пенициллин G |
>100 |
|
Ампициллин |
>100 |
|
Тетрациклин |
0,05-1,0 |
|
Доксициклин |
0,06-0,25 |
|
Миноциклин |
0,016-0,063 |
|
Эритромицин |
0,01-1,0 |
|
Азитромицин |
0,06-0,5 |
|
Кларитромицин |
0,004-1,0 |
|
Офлоксацин |
0,125-2,0 |
|
Ципрофлоксацин |
16,0 |
|
Спарфлоксацин |
<0,5 |
|
Грепафлоксацин |
<0,5 |
|
Левофпоксацин |
0,125-0,5 |
|
Моксифлоксацин |
<0,5 |
|
Тровафлоксацин |
<0,5 |
|
Гемифлоксацин |
<0,25 |
|
Троспектиномицин |
>400 |
|
Супьфисоксазол |
10-20 |
In vitro С. pneumonia обладает резистентностью к ряду представителей беталактамных антибиотиков (пенициллины и цефалоспорины) [98]. Возбудитель чувствителен к эритромицину, кларитромицину, азитромицину (МПК90 < 0,5 мкг/мл) [99, 100]. Новый представитель макролидов HMR 3647 in vitro обладает высокой чувствительностью против штаммов С. pneumonia. Его МПК90 составила 0,25 мкг/мл (диапазон от 0,15 до 2 мкг/мл) [98,101-106].
Штаммы С. pneumonia чувствительны к ряду представителей новейших фторхинолоновых препаратов IV поколения: грепафлоксацину, левофлоксацину, моксифлоксацину, тровафлоксацину и др. (МПК90 < 0,5 мкг/мл). Наиболее активным из них является гемифлоксацин (МПК90 < 0,25 мг/мл) [100, 107, 108].
Исследование минимальных подавляющих концентраций (МПК) нового фторхинолонового препарата АМ-1155 в отношении штаммов С. pneumonia показало, что диапазон МПК90 составил от 0,063 до 0,125 мкг/мл. Данный препарат обладает активностью, схожей с спарфлоксацином и тосуфлоксацином. В то же время она была выше активности левофлоксацина и ципрофлоксацина в 4 и 16 раз соответственно, но ниже, чем кларитромицина и миноциклина (диапазон МПК от 0,016 до 0,031 и от 0,016 до 0,063 мкг/мл соответственно) [109].
Макролиды, рокситромицин, азитромицин, кларитромицин, доксициклин и миноциклин, а также некоторые представители фторхинолоновых препаратов III и IV поколений являются потеннциальными антихламидийными препаратами, которые могут использоваться в случаях неосложненной инфекции, вызванной С. pneumonia. Отмечено, что не всегда после проведенного стандартного курса (7-10 дней) антимикробной терапии при остром респираторном заболевании, вызванном С. pneumonia, происходит эрадикация возбудителя. При пневмониях хламидийной этиологии высокой эффективностью обладают новые макролиды и фторхинолоны. Например, тровафлоксацин обладает большей эффективностью в лечении инфекций дыхательных пуши, вызванных хламидиями, по сравнению с офлоксацином и ципрофлоксацином [ПО]. В то же время имеются данные о полном излечении больных с хламидийной пневмонией при использовании офлоксацина [97],
В сравнительных исследованиях было показано, что азитромицин более эффективен, чем эритромицин, в лечении атипичных пневмоний. Данные препараты являются основой лечения хламидийных и микоплазменной пневмоний. Азитромицин при однократном применении в течение 5 дней оказывает терапевтический эффект, схожий с использованием амоксициллинклавулоновой кислоты и эритромицина, назначаемыми три раза в день в течение 10 дней. Кроме того, данный препарат обладает меньшими побочными эффектами [111], и обычный курс химиотерапии при инфекции дыхательных путей составляет, по данным различных авторов, 5-10-14 дней при суточной дозировке 1,5 г. Тетрациклиновые антибиотики относят к группе альтернативных препаратов при пневмониях, вызванных данным возбудителем, и назначаются в течение 2-3 недель [112]. Эффективность лечения пневмоний, вызванных С. pneumonia, моксифлоксацином (новый хинолоновый препарат) у взрослых оценивают в 70%, кларитромицином – 79%, азитромицином – до 83% и эритромицином – 86%.
Имеются данные об успешном лечении астмы после инфекции С. pneumonia у взрослых при проведении месячного курса терапии макролидами или антибиотиками тетрациклинового ряда. 6-12-недельный курс печения этими препаратами рекомендован при реактивных артритах хпамидийной этиологии.
Ранее уже сообщалось, что С. pneumonia и специфические Т-лимфоциты С. pneumonia часто присутствуют в стенке аневризмы брюшной аорты, что подтверждает возможную роль этого микроорганизма в патогенезе данного заболевания. Исследования на 36 больных с аневризмой брюшной аорты показали, что использование антимикробной терапии (доксициклин 150 мг/сутки), направленной на элиминацию возбудителя, уменьшает рост аневризмы. Эти результаты подтверждены ультрасонографией при наблюдении больных в течение 18 месяцев в группах, получавших антибиотик (19 человек) и плацебо (17 человек) [113].
Изучение влияния антибактериальных препаратов на течение инфаркта миокарда и атеросклероза, кофактором которых, возможно, является С. pneumonia, требует всесторонних и углубленных исследований. По-видимому, в таких случаях будет необходима продолжительная терапия в течение 6-12 недель [46]. В то же время при хроническом процессе, особенно если речь идет об атеросклерозе или астме и наличии персистирующей инфекции С. pneumonia, длительность терапии установить довольно сложно, так как неизвестно, как долго могут выявляться в тканях организма ДНК от погибших хламидий (мишень для ПЦР-анализа) или антигены С. pneumonia (мишень для ICC). Вследствие этого трудно установить длительность курсов лечения азитромицином, которое приводило бы к эрадикации живых клеток С. pneumonia из ткани легкого или пораженных сосудов сердца. По-видимому, длительные курсы (4-6 недель) антибиотикотерапии необходимы при инфицировании С. pneumonia глубоких тканей и органов. Так как культуру С. pneumonia изолировать при наличии хронической инфекции очень трудно, то в качестве теста эффективности лечения вызываемой ею инфекции возможно использование метода RT-ПЦР-анализа [114]. Проведенные в ФРГ исследования на людях в возрасте 50-69 лет показали, что по сравнению с пенициллином макропиды имеют возможный профилактический эффект в развитии кардиоваскулярных симптомов на протяжении первых шести месяцев [115].
Специфическая профилактика
Специфическая профилактика отсутствует. В настоящее время проводятся исследования по разработке ДНК-вакцин, содержащих гены, кодирующие МОМР, отр-2 и Hsp 60, которые в экспериментах на животных показали обнадеживающие результаты, свидетельствующие о возможности создания вакцины против С. pneumonia [116, 117].
Заключение
Рассмотренный патоген С. pneumonia является причиной развития разнообразных патологических состояний у человека, который на протяжении жизни может часто подвергаться реинфекции данным микроорганизмом.
Сведения о заболеваемости людей, вызванной Chlamydia pneumonia, в России практически отсутствуют в отечественной литературе. Отсутствуют и сертифицированные тест-системы дня диагностики данного возбудителя. Не разработаны также средства специфической профилактики заболеваний, вызываемых С. pneumonia. Авторы надеются, что представленные в настоящем обзоре сведения привлекут внимание клиницистов к многообразной патологии, вызываемой данным возбудителем, и окажутся полезными для специалистов, разрабатывающих средства диагностики, лечения и профилактики инфекций, вызванных С. pneumonia.
Литература
1. Everett К. D. E, Bush R. В., Andersen A. A. Emended description of the order Chlamydiales, proposal of Parachlamydiaceae fam. nov. and Simkaniaceae fam. nov., each containing one monotypic genus, revised taxonomy of the family Chlamydiaceae, including a new genus and five species, and standards for the identification organisms // International Journal of Systematic bacteriology. 1999. 49. P. 415-440.
2. Rurangirwa F. R., Dilbeck P. M., Crawford Т. В., McGuire T. C., McElwain T. F. Analysis of the 16S rRNA gene of micro-organism WSU 86-1044 from an aborted bovine foetus reveals that it is a member of the order Chlamydiales: proposal of Waddliaceae fam. nov., Waddlia chondrophila gen. nov., sp. nov. // Int. J. Syst. Bacteriol. 1999. Apr. 49. P. 577-81
3. Intracellular bacterial Infections / Ed. J. C. Pechere. 1996. P. 53-59.
4. Read T. D., Brunham R. C., Eisen J., Fraser C. M. The genomics of chlamydia species // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. Helsinki, Finland, 20-23 August 2000. P. 29.
5. Wuppermann F. N., Schwab J. et al. Physical and genetic mapping of the chlamydia pneumonia genome // Abstracts of Proceedings of the 3rd Meeting of the European Society for Chlamydial Research. Vienna, Austria, 11-14 September 1996. P. 42.
6. Thomas N. S., Storey C. et al. The cryptic plasmid of Chlamydia pneumoniae: complete nucleotide sequence reveals a deletion in ORF1 // Ibid. P. 44.
7. Wiedmann-AI-Ahmad M., Schon G., Freidank H. Protein patterns of Chlamydia pneumonia strains and immunoblot analysis // Ibid. P. 56.
8. Knudsen K., Madsen A. S. et al. Differences in the outer membrane complex between Chlamydia pneumonia and chlamydia trachomatis // Ibid. P. 17.
9. Mygind P., Birkelund and Christiansen. Prevalence of antibodies to Chlamydia outer membrane protein 2, GroEL and DnaK in suspected acute C. pneumonia Infections // Ibid.
10. Егоров A. M., Сазыкин Ю. О. Хламидии: Молекулярная организация клетки и некоторые особенности патогенеза инфекций // Антибиотики и химиотерапия. 2000. 45. 4. С. 3-5.
11. Schuhmacher A., Melchers К. et al. The type III secretion system of Chlamydia pneumoniae // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 47.
12. Friedman M. G., Birkeback N. et al. Serologic evidence for infection with Simkania negevensis in Denmark and its possible assaciation with chronic cough // Ibid. P. 258.
13. Grayston J. T. Epidemiology of Chlamydia pneumonia (TWAR) // Abstracts of Proceedings of the 3rd Meeting of the European Society for Chlamydial Research. P. 2111-212.
14. Kauppinen M., Saikku P. Pneumonia due to Chlamydia pneuminiae: prevalence, clinical features, diagnosis and treatment // Clinical Infectious Diseases. 1995. 21. P. 244-252.
15. Block S. L., Hammerschlag M. R., Hedrick J. et al. Chlamydia pneumoniae in acute otitis media // Pediatr. Infect. Dis. J. 1997. Sep. 16(9). P 858-62.
16. Dake Y., Enomoto Т., Shibano A., Sakoda Т., Saito Y., Takahashi M. Epidemiologic study of Chlamydia pneumoniae with ELISA // Nippon Jibiinkoka Gakkai Kaiho. 1998. Nov. 101 (11). P. 1316-20.
17. Saikku P. Chronic Chlamydia pneumoniae infections //Abstracts of Proceedings of the 3rd Meeting of the European Society for Chlamydial Research. P. 215-218.
18. Wong Y. K., Gallagher P. J., Wardc M. E. Chlamydia pneumonia and atherosclerosis // Heart. 1999. 81. P. 232-238.
19. Quinn T. C., Gaydos C. A. In vitro infection and pathogenesis of Chlamydia pneumoniae in endovascular cells // Am. Heart J. 1999. Nov. 138 (5 Pt 2). P. 507-511.
20. Redecke V., Dalhoff K., Bohnet S., Braun J., Maass M. Interaction of Chlamydia pneumoniae and human alveolar macrophages: infection and inflammatory response // Am. J. Respir. Cell. Mol. Biol. 1998. Nov. 19(5). P. 721-7.
21. Hahn D. L., McDonald R. Can acute Chlamydia pneumoniae respiratory tract infection initiate chronic asthma? // Ann. Allergy Asthma Immunol. 1998. Oct. 81(4). P. 339-44.
22. Ramos P., Arrictu L., Samaniego S. et al. Chlamydia pneumoniae infection in astma and coronary heart patients: serology study // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 135.
23. Hahn D. L. Chlamydia pneumoniae, asthma, and COPD: what is the evidence? // Ann. Allergy Asthma Immunol. 1999. Oct. 83(4). P. 271-88, _291, 291-2.
24. Kleemola M., Saikku P., Visakorpi R., Wang S. P., Grayston J. T. Epidemics of pneumoniae caused by TWAR, anew Chlamydia organ-ism, in military trainees in Finland // J. Infect. Dis. 1988. 157. P. 230-236.
25. Comandini U. V., Maggi P., Santopadre P., Monno R., Angarano G., Vullo V. Chlamydia pneumoniae respiratory infections among patients infected with the human immunodeficiency virus // Eur. J. Clin. Microbiol. Lnfect. Dis. 1997. Oct. 16(10). P. 720-6.
26. Koyi H., Branden E., Gnarpe J. et al. Chlamydia pneumoniae may be associated with lung cancer: Preliminary report on a seroepidemiological study // APMIS. 1999. Sep. 107(90). P. 828-32.
27. Gencay M., Tamm M. et al. Chronic C. pneumoniae infection is assaciated with epithelial cell carcinoma // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial_Research. P. 259.
28. Danesh J., Collins R., Peto R. Chronic infections and coronary heart disease: is there a link? // Lancet. 1997. Vol. 350. P. 430-6.
29. Laurila A., Bloigu A., Nayha S. et al. Chronic Chlamydia pneumoniae infection is associated a serum lipid profile known to be a risk factor for atherosclerosis // Arterioscler. Thromb. Vase. Biol. 1997. Vol. 17. P 2910-13.
30. Ridker P. M., Kundsin R. В., Stampfer M. J. et al. Prospective study of Chlamydia pneumoniae IgG seropositivity and risks offuture myocardial infarction // Circulation, 1999. Vol. 99, P. 1161-64.
31. Saikku P., Leinonen M., Manila K. et al. Serologic evidence of an association of a novel Chlamydia, TWAR, with chronic coronary heart disease and acute myocardial infarction // Lancet. 1988. 2. P. 983-6.
32. Saikku P., Leinonen M., Tenkanen L. et al. Chronic Chlamydia pneumoniae infection as a risk factor for coronary heart disease in the Helsinki Heart Study //Ann. Intern Med. 1992. Vol. 116, P, 273-8.
33. Meniconi A., Noll G., Luscher T. F. Is arteriosclerosis an infectious disease? // Schweiz. Rundsch. Med. Prax. 1998. Vol. 87. P. 64-74.
34. Nishimura M., Ushiyama M., Nanbu A., Mashida C., Kawagoe K., Yoshimura M. Inverse association of Chlamydia pneumoniae infection with high blood pressure in Japanese adults // Am. J. Hypertens. 2001. Jan. 14(1). P. 20-6.
35. Gabriel A. S., Gnarpe H., Hallander H. et al. The prevalence of chronic Chlamydia pneumoniae infection as detected by polymerase chain reaction in pharyngeal samples from patients with ischaemic heart disease // Europ. Heart J. 1998. Vol. 19, P 1321-7.
36. Grayston J. T. Chlamydia pneumoniae, strain TWAR pneumoniae // Annu. Rev. Med. 1992. 43. P. 317-23.
37. Leinonen M., Saikku P. Interaction of Chlamydia pneumoniae infection with other risk factors of atherosclerosis // Am. Heart J. 1999. Nov. 138(5 Pt 2). P. 504-6.
38. Glader C. A., Stegmayr B. et al. Chlamydia pneumoniae Antibodies and High Lipoprotein(a) Levels Do Not Predict Ischemic Cerebral Infarctions: Results From a Nested Case-Control Study in Northern Sweden // Stroke. 1999. Oct. 30(10). P. 2013-18.
39. Murray L. J., O”Reilly D. P., Ong G. M., O”Neill C., Evans A. E., Bamford K. B. Chlamydia pneumoniae antibodies are associated with an atherogenic lipid profi1e// Heart. J. 999. Mar 81(3). P. 239-44.
40. Thorn D. H., Wang S. P., Grayston J. T. et al. Chlamydia pneumoniae strain TWAR antibody and angiographically demonstrated coronary artery disease //Arterioscler. J Thromb. 1991. Vol. 11. P. 547-51.
41. Bachmaier K., Neu N., de la Maza L. M., Pal S., Hessel A., Penninger J. M. Chlamydia Infections and Heart Disease Linked Through Antigenic Mimicry // Science. 1999. Feb. 26. 283 (5406). P. 1335-9.
42. Tiran В., Tiesenhausen K., Karpf E. et al. LPS IgG antibodies are a marker for endovascular presence of Chlamydia pneumoniae in the atherosclerotic carotid artery // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 295.
43. Weib В., Wuppermann F. N. et al. Chlamydia pneumonia infection alters gene expression in endotelial cells // Ibid. P. 297.
44. Anttila Т., Leinonen M., Surcel H. M., Von Hertzen L., Bloigu A., Seppala I., Saikku P. IgG subclass-specific antibodies in Chlamydia pneumoniae infections // Scand. J. Infect. Dis. 1998. 30 (4). P. 381-6.
45. Gurfinkel E. P., Bozovich G. Emerging role of antibiotics in atherosclerosis // Am. Heart J. 1999. Nov. 138(5 Pt 2). P. 537-8.
46. Stille W., Dittmann R. Arteriosclerosis as a sequela of chronic Chlamydja pneumoniae infection // Herz. 1998. May 23(3). P. 185-92.
47. Meier C., Derby L., Jick S. et al. Antibiotics and risk of subsequent first-time acute myocardial infarction // JAMA. 1999. 281. P. 427-31.
48. Martin D., Bursill J., Qui M. R. et al. Alternative hypothesis for efficacy of macrolides in acute coronary syndromes // Lancet. 1998. 351. P. 1858-9.
49. Gupta S., Leatham E. W., Carrington D. et al. Elevated Chlamydia pneumoniae antibodies, cardiovascular events, and azithromycin in male survivors of myocardial infarction // Circulation. 1997. Vol. 96. P. 404-7.
50. Gurfinkel E., Bozovich G., Beck E. et al., for the ROXIS Study Group. Treatment with the antibiotic roxithromycin in patients with acute non-Q-wave coronary syndromes: The final report of the ROXIS study // Europ. Heart J. 1999. Vol. 20. P. 121-7.
51. Folsom A. Antibiotics for prevention of myocardial infarction? Not yet! // JAMA. l999. 281. P. 461-2.
52. Weiss S. M., Roblin P. M., Gaydos C. A. et al. Failure to detect Chlamydia pneumoniae in coronary atheromas of patients undergoing atherectomy // J. Infect. Dis. 1996. Vol. 173. P. 957-62.
53. Libby P., Egan D., Skarlatos S. Roles of infectious agents in atherosclerosis and restenosis. // Circulation. 1997. 96. P. 409-10.
54. Saikku P. Epidemiology of Chlamydia pneumoniae in atherosclerosis // Am. Heart J. 1999. Nov. 138(5 Pt 2). P. 500-3.
55. Grayston J. T. Chlamydia pneumoniae (TWAR) and coronary heart disease: from hypothesis to treatment // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 380-381.
56. Dean D., Molestina R. et al. Molecular and biological characterization of a unique isolate of chlamydia pneumonia from a coronary atheroma // Abstracts of Proceedings of the 3rd Meeting of the European Society for Chlamydial Research. P. 43.
57. Sessa R., Pietro M. D., Schiavoni G. et al. Prevalence of Chlamydia pneumoniae DNA in atherosclerotic plaques of carotid arteries // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 288.
58. Byrne G. I., Kalayoglu M. V. Chlamydia pneumoniae and atherosclerosis: Links to the disease process // Am. Heart J. 1999. Nov. 138 (5 Pt 2). P. 488-90.
59. Vehmaan-Kreula P., Puolakkainen M., Sarvas M., Welgus H. G., Kovanen P. T. Chlamydia pneumoniae proteins induce secretion of the 92-kDa gelatinase by human monocyte-derived macrophages // Arterioscler. Thromb. Vase. Biol. 2001. Jan. 21(1). P. E1-8.
60. Grayston J. T, Kuo C. C., Coulson A. S. et al. Chlamydia pneumoniae (TWAR) in atherosclerosis of the carotid artery // Circulation. 1995. Vol. _92. P. 3397-3400.
61. Coombes В. К., Mahony J. В. cDNA array analysis of altered gene expression in human endothelial cells in response to Chlamydia pneumoniae infection // Infect. Immun. 2001. Mar 69(3). P. 1420-7.
62. Dunne M. WIZARD and the design of trials for secondary prevention ofatherosclerosis with antibiotics // Am. Heart J. 1999. Nov. 138(5 Pt 2). P. 542-4.
63. Fong I. W., Chiu В., Viira E., Jang D., Mahony J. B. De novo induction of atherosclerosis by chlamydia pneumoniae in a rabbit model // Infect. Immun. 1999. Nov 67(11). P. 6048-55.
64. Laitinen K., Laurila A., Pyhala L., Leinonen M., Saikku P. Chlamydia pneumoniae infection induces inflammatory changes in the aortas of rabbits // Infect. Immun. 1997. Nov 65(11). P. 4832-5.
65. Airenne S., Surcel H. M., Alakarppa H., Laitinen K., Paavonen J., Saikku P., Laurila A. Chlamydia pneumoniae infection in human monocytes // Infect. Immun. Mar 67(3). P. 1445-9.
66. Smiega M., Chong S., Natarajan M. et al. Circulating Chlamydia pneumoniae DNA is associated with smoking and season in coronary angiography patients // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 291.
67. Blessing E., Campbell L. A., Rosenfeld M., Kuo С. С. Infection with chlamydia pneumoniae accelerates the induction and progression of atherosclerosis in diet induced hypercholesteremic С 57 BL/6S mice. P. 247.
68. Liu L, Hu H., Ji H., Murdin A. D., Pierce G. N., Zhong G. Chlamydia pneumoniae infection significantly exacerbates aortic atherosclerosis in an LDLR-/- mouse model within six months // Mol. Cell. Biochem. 2000. Dec. 215(1-2). P. 123-8.
69. Rothstein N. M., Quinn T. С., Madico G., Gaydos С. A., Lowenstein С. J. Effect of Azithromycin on Murine Arteriosclerosis Exacerbated by Chlamydia pneumoniae // J. Infect. Dis. 2001. Jan. 183(2). P. 232-8.
70. Chiu B. Multiple infections in carotid atherosclerotic plaques // Am. Heart J. 1999. Nov. 138 (5 Pt 2). P. 534-6.
71. Coles K. A., Plant A. J., Riley T. V., Smith D. W., McQuillan B. M., Thompson P. L. Lack of association between seropositivity to Chlamydia pneumoniae and carotid atherosclerosis // Am. J. Cardiol. I999. Oct. 1. 84(7). P. 825-8.
72. Valtonen V. V. Role of infections in atherosclerosis // Am. Heart J. 1999. Nov. 138 (5 Pt 2). P. 825-8.
73. Maraha В., den Heijer M., Wullink M., van der Zee A. et al. Detection of Chlamydia pneumoniae DNA in buffy-coat samples of patients with abdominal aortic aneurysm // Eur. J. Clin. Microbiol. Infect. Dis. 2001. Feb. 20(2). P. 111-6.
74. Karlsson L, Gnarpe J., Naas J. et al. Detection of viable Chlamydia pneumoniae in abdominal aortic aneurysms // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 266.
75. Blasi F, Denti F, Erba M. et al. Detection of Chlamydia pneumoniae but not Helicobacter pylori in atherosclerotic plaques of aortic aneurisms // J. Clin. Microbiology. 1996. Vol. 34. P. 2766-9.
76. Wimmer M. L. J., Hobert R. L Chlamydia pneumoniae and Neurological Disorders // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 261-2.
77. Balin В. J., Gerard H. C., Arking E. J., Appelt D. M., Branigan P. J., Abrams J. Т., Whittum-Hudson J. A., Hudson A. P. Identification and localization of Chlamydia pneumoniae in the Alzheimer”s Brain // Med. Microbiol. Immunol. (Berl.). 1998. Jun. 187(1). P. 23-42.
78. Ossewaarde J. M., Gielis-Proper S. K., Meijer A., Roholl P. J. M. Chlamydia pneumoniae antigenes are present in the brains of Alzheimer patients, but not in the brains of patients with other dementia”s // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 284.
79. Machony J., Wouife J., Munoz D. et al. Chlamydia pneumoniae in the alzheimer”s brain – is DNA detection hampered by low copy number? // Ibid.
80. Sriram S., Stratton C. W., Yao S., Tharp A., Ding L., Bannan J. D., Mitchell W. M. Chlamydia pneumoniae infection of the central nervous system in multiple sclerosis // Ann. Neurol. 1999. Jul. 46(1). P. 6-14.
81. Zuzak К. В., Theodore M., Kayfman M. et al. Lack of detection of Chlamydia pneumoniae by PCR and tissue culture in cerebrospinal fluids from multiple sclerosis patients and controls // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 303.
82. Marcos M. A., Sanz A., Vidal J. Lack of detection of Chlamydia pneumoniae in cereprospinal fluid from patients with multiple sclerosis // Ibid. P. 278.
83. Miyashita N., Saida Т., Matsui M. et al. Does Chlamydia pneumoniae play a role in pathogenesis of multiple sclerosis // lbid. P. 280.
84. Paavonen J. Chlamydia and cancer // Ibid. P. 239-40.
85. Otzipka E., Fenner Th. et al. Chlamydia pneumoniae and lymphadenitis: Possibility of early diagnosis – follow up by defined Chlamydia rLPS-Serology: A case report // Abstracts of Proceedings of the 3rd Meeting of the European Society for Chlamydial Research. P. 231.
86. Kanauchi M., Kawano Т., Dohi К. Association of Chlamydia pneumoni-ae infection with diabetic nephropathy // Diabetes Res. Clin. Pract. 2000 Jan. 47(1). P. 45-8.
87. Schumacher H. Ft., Jr, Gerard H. C. et al. Lower prevalence of ff Chlamydia pneumoniae DNA compared with Chlamydia trachomatis 1-p DNA in synovial tissue of arthritis patients // Arthritis Rheum. 1999. Sep. 42(9). P. 1889-93.
88. Schumacher R. H., Jr. Chlamydial artritis // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 229-30.
89. Boman J. Diagnosis of Chlamydia pneumoniae infections // Ibid. P. 65-58.
90. Naas J., Gnarpe J. A. Demonstration of Chlamydia pneumoniae in tissue by immunohistoctemistry // APMIS. 1999. Sep 107(9). P. 882-6.
91. Anttila T, Leinonen M., Surcel H. M., von Hertzen L. et al. IgG subclass-specific antibodies in Chlamydia pneumoniae infections // Scand. J. Infect. Dis. 1998. 30 (4). P. 381-6.
92. Saikku P. Diagnosis of acute and chronic Chlamydia pneumoniae infections // Abstract Proceeding of the Eighth International Symposium on Human Chlamydia Infections, 19-24 June 1994. P. 163-72.
93. Farholt S., Boman J. et al. Chlamydia pneumoniae PCR: quality control is needed. P. 95.
94. Apfalter P., Blasi F., Boman J., Gaydos C. A. et al. Multicenter Comparison Trial of DNA Extraction Methods and PCR: Assays for Detection of Chlamydia pneumoniae in Endarterectomy Specimens // J. Clin. Microbiol. 2001. Feb. 39(2). P. 519-24.
95. Apfalter P., Durack D. T. and Gaydos C. A. Detection of Chlamydia pneumoniae by real-time strand displacment // Abstracts of Proceedings of the IV Meeting of the European Society for Chlamydial Research. P. 99.
96. Gnarpe J., Lindquist L. A new DIG-PCR-EIA method for the detection of Chlamydia pneumoniae DNA in clinical samples // APMIS. 2000. Sep. 108(9). P. 626-32.
97. Манзенюк И. H., Воробьева M. С., Ямникова С. С. Пневмония, вызванная Chlamydophila (Chlamydia) pneumoniae: клиника, диагностика и лечение // Антибиотики и химиотерапия. 2001. 46. 1. С. 22-29.
98. Kuo С. С. and Grayston J. T. In vitro drug susceptibility of Chlamydia sp. Strain TWAR // Antimicrob. Agents Chemother. 1988. 32. P. 257.
99. Roblin P. M., Hammerschlag M. R. Microbiologic efficacy of azithromycin and susceptibilities to azithromycin of isolates of Chlamydia pneumoniae from adults and children with community-acquired pneumonia // Antimicrob. Agents Chemother. 1998. Jan. 42(1). P. 194-6.
100. Roblin P. M., Kutlin A. et al. Activity of grepafloxacin and other fluoroquinolones and newer macrolides recent clinical isolates of Chlamydia pneumonia // J. Antimicrob. Agents. 1999. Jul. 12(2). P. 181-4.
101. Roblin P. M., Hammerschlag M. R. In vitro activity of a new ketolide antibiotic, HMR 3647, against Chlamydia pneumoniae // Antimicrob. Agents Chemother. 1998. Jun. 42(6). P. 1515-6.
102. Cooper M. A., Baldwin D., Matthews R. S. et al. In vitro susceptibility of Chlamydia pneumoniae (TWAR) to seven antibiotics // J. Antimicrob. Chemother. 1991. 28. P. 407.
103. Hammerschlag M. R., Qumei K. K. and Roblin P. M. In vitro activity of azithromycin, clarithromycin, L-ofloxacin and other antibiotics against Chlamydia pneumoniae // Antimicrob. Agents Chemother. 1992. 36. P. 1573.
104. Welsh L. E., Gaydos C. A. and Quinn Т. С. In vitro evaluation of activities of azithromycin, erythromycin and tetracycline against Chlamydia trachomatis and Chlamydia pneumoniae // Ibid. P. 291.
105. Id. In vitro activities of azithromycin, clarithromycin, erythromycin and tetracycline against 13 strains of Chlamydia pneumoniae // Antimicrob. Agents Chemother. 1996. 40. P. 212-14.
106. Gnarpe J., Eriksson K., Gnarpe H. In vitro activities of azitromycin and doxycicline against 15 isolates of Chlamydia pneumoniae // Ibid. P. 184.
107. Roblin P. M., Reznik T, Kutlin A., Hammerschlag M. R. In Vitro Activities of Gemifloxacin (SB 265805, LB20304) against Recent Clinica Isolates of Chlamydia pneumoniae // Antimicrob. Agents Chemother. 1999. Nov. 43(11). P. 2806-7.
108. Roblin P. M., Hammerschlag M. R. In vitro activity of GAR-936 against Chlamydia pneumoniae and Chlamydia trachomatis // Int. J. Antimicrob. Agents. 2000. Sep. 16(1). P. 61-3.
109. Miyashita N, Niki Y, Kishimoto T, Nakajima M, Matsushima T. In vitro and in vivo activities of AM-1155, a new fluoroquinolone, against Chlamydia spp. // Antimicrob Agents Chemother. 1997. Jun. 41(6). P. 1331-4.
110. Pechere J. C., Gootz T. D. Bacteriological activity of trovafloxacin, a new quinolone, against respiratory tract pathogens // Eur. J. Clin. Microbiol. Infect. Dis. 1998. 17 (6). P. 405-12.
111. Harris J. A., Kolokathis A., Campbell M., Cassell G. H., Hammerschlag M. R. Safety and efficacy of azithromycin in the treatment of community-acquired pneumonia in children // Pediatr. Infect. Dis. J. 1998. Oct. _ 17(10). P. 865-71.
112. Hammerschlag M. R. Antimicrobial susceptibility and therapy of infections caused by Chlamydia pneumoniae // Antimicrob. Agents Chemother. 1994. J38_P. J 873-8.
113. Mosorin M., Juvonen J., Surcel H. M. et al. Doxicyclin decreases the growth rate of abdominal aortic aneureysm prospective, randomized, double blinded study // Abstracts of Proceedings of the IV Meeting of _the European Society for Chlamydial Research. P. 399.
114. Hang D., Cambell L. A., Kuo С. С. Failure of four and six weeks of treatment to eradicate evidence of Chlamydia pneumoniae from human lung and vascular tissue: pathology case reports // Ibid. P. 395.
115. Ostergaard L., Sorensen H. Т., Lindholt J. et.al. Risk of cardio-vascular events in patients aged 50-70 years treated with macrolides versus penicillins // Ibid. P. 421.
116. Pentila Т., Vuola J. M., Puurala V. et al. Immunity to Chlamydia pneumoniae induced by DNA vaccination // Ibid. P. 403.
117. Penttila Т., Vuola J. M., Puurula V, Anttila M., San/as M., Rautonen N., Makela P. H., Puolakkainen M. Immunity to Chlamydia pneumoniae induced by vaccination with DNA vectors expressing a cytoplasmic protein (Hsp60) or outer membrane proteins // Vaccine. 2000. Dec. 8. 19(9-10). P. 1256-65.
Anti-Chlamydophila pneumonia-IgG (антитела класса IgG к Chlamydophila pneumonia)
Метод определения Иммунохимический (иммуноферментный) анализ.
Исследуемый материал Сыворотка крови
Доступен выезд на дом
Онлайн-регистрацияМаркер текущей или имевшей место в прошлом инфекции Chlamydia pneumoniae.
Chlamydia pneumoniae, подобно Chlamydia trachomatis, является бактерией – облигатным внутриклеточным паразитом. Это широко распространенный патоген, передающийся от человека к человеку респираторным путем. Высокая частота инфекции Chlamydia pneumoniae отмечается среди детей школьного и дошкольного возраста. Антитела к данному патогену выявляются у более 50% взрослого населения.
Клинические проявления инфекции Chlamydia pneumoniae связывают в основном с такими патологическими состояниями, как хронические инфекции верхних дыхательных путей, бронхиты, пневмонии, и их осложнениями. Результаты клинических исследований последних лет говорят о вероятности связи инфекции Chlamydia pneumonia и воспалительного ответа организма на нее с механизмами развития астмы и атеросклероза.
В связи со сложностью выделения Chlamydia pneumonia, в лабораторной диагностике данной инфекции используется ПЦР-исследования, а также находят применение серологические тесты (определение IgM, IgA, IgG антител в сыворотке крови). Инфекция Chlamydia pneumoniae развивается медленно, инкубационный период составляет несколько недель – больше, чем для многих других респираторных инфекций. IgM антитела появляются примерно через 3 недели после начала заболевания в невысоких титрах и начинают снижаться через 2 месяца. IgG и IgA могут не выявляться до 6 – 8 недель от начала заболевания, затем титры могут быстро расти. достигая высоких уровней. IgA имеют короткий период полувыведения, IgG могут выявляться до 3 лет.
Стойкого иммунитета к Chlamydia pneumoniae не возникает, случаи повторного инфицирования являются обычными. Свидетельством текущей или недавней инфекции является наличие IgM антител и 3 – 4 кратный рост титров IgA и IgG в течение 1 – 2 недель. Вследствие высокой частоты выявления антител среди населения, серопозитивность не является адекватным маркером хронической инфекции, хотя длительное сохранение высоких титров IgG антител и персистенция IgA антител может свидетельствовать о хронической инфекции Chlamydia pneumoniae.
Литература
- Руководство по инфекционным болезням. Под редакцией: Ю.В. Лобзина, С.С. Козлова А.Н. Ускова. Спб., Феникс, 2001, 932 с.
- Tiina Sävykoski née Huittinen. Chlamydia pneumoniae infection, inflammation and heat shock protein 60 immunity in asthma and coronary heart disease. Acad. Diss. . University of Oulu, 2003. http://herkules.oulu.fi/isbn9514269853.
- Schneeberger PM, Dorigo-Zetsma JW, van der Zee A, et al: Diagnosis of atypical pathogens in patients hospitalized with community-acquired respiratory infection. Scand J Infect Dis 2004, v. 36(4), pp. 269 – 73.
- Gorbach S. Et al./ Infectious Diseases (3rd edition)/2003/ Lippincott Williams & Wilkins/2700 ps.
Определение антител классов M (IgM) к хламидии пневмонии (Chlamydia pneumoniae) в крови
АНМО «Ставропольский краевой клинический консультативно-диагностический центр»:
355017, г. Ставрополь, ул. Ленина 304(8652) 951-951, (8652) 35-61-49 (факс)
(8652) 951-951, (8652) 31-51-51 (справочная служба)
Посмотреть подробнееОбособленное подразделение «Диагностический центр на Западном обходе»:
355029 г. Ставрополь, ул. Западный обход, 64(8652) 951-951, (8652) 31-51-51 (контактный телефон)
(8652) 31-68-89 (факс)
Посмотреть подробнееКлиника семейного врача:
355017 г. Ставрополь, пр. К. Маркса, 110 (за ЦУМом)(8652) 951-951, (8652) 31-51-51 (контактный телефон)
(8652) 31-50-60 (регистратура)
Посмотреть подробнееНевинномысский филиал:
357107, г. Невинномысск, ул. Низяева 1(86554) 95-777, 8-962-400-57-10 (регистратура)
Посмотреть подробнееОбособленное структурное подразделение в г. Черкесске :
369000, г. Черкесск, ул. Умара Алиева 318(8782) 26-48-02, +7-988-700-81-06 (контактные телефоны)
Посмотреть подробнееОбособленное структурное подразделение в г. Элисте :
358000, г. Элиста, ул. Республиканская, 478(989) 735-42-07 (контактные телефоны)
Посмотреть подробнееЗАО “Краевой клинический диагностический центр”:
355017 г. Ставрополь, ул. Ленина 304(8652) 951-951, (8652) 35-61-49 (факс)
(8652) 951-951, (8652) 31-51-51 (справочная служба)
Посмотреть подробнееОбособленное структурное подразделение на ул. Савченко, 38 корп. 9:
355021, г. Ставрополь, ул. Савченко, 38, корп. 98 (8652) 316-847 (контактный телефон)
Посмотреть подробнееОбособленное структурное подразделение на ул. Чехова, 77 :
355000, г. Ставрополь, ул. Чехова, 778(8652) 951-943 (контактный телефон)
Посмотреть подробнееОбособленное структурное подразделение в г. Михайловске:
358000, г. Михайловск, ул. Ленина, 201 (в новом жилом районе «Акварель»).8(988) 099-15-55 (контактный телефон)
Посмотреть подробнееАнтитела к Chlamydia pneumoniae, IgG
Антитела к Chlamydia pneumoniae, IgG
Chlamydia (Chlamydophila) pneumoniae – внутриклеточный паразитарный микроорганизм, источник инфекционного поражения органов дыхания. Он не способен самостоятельно синтезировать АТФ и другие метаболические ресурсы энергии, поэтому может существовать только внутри клеток. C. pneumoniaе вызывает поражения верхних дыхательных путей (ларингит, фарингит, синусит, средний отит, бронхит) и пневмонию у детей и молодежи, а также рецидивирующие инфекционные заболевания органов дыхания у старших возрастных групп. C. pneumoniaе передается только от человека к человеку. Вспышки респираторных заболеваний хламидийной этиологии чаще бывают в тесных коллективах. У 70 % зараженных инфекция может бессимптомно персистировать в дыхательных органах. Инкубационный период точно не установлен. Иногда заболевание проявляется через много месяцев после инфицирования, а бактерии способны длительно сохраняться на слизистых верхних дыхательных путей даже после выздоровления человека. Воспаление легких, вызванное C. Pneumoniae (по некоторым данным, это около 10 % всех случаев), относится к атипичным пневмониям. У них нет специфических симптомов, и течение заболевания похоже на микоплазменную инфекцию. В ответ на инфицирование Chlamydia) pneumoniae иммунная система вырабатывает специфические иммуноглобулины: IgA, IgM и IgG.
Иммуноглобулины IgG вырабатываются в ответ на антигены C. pneumoniaе через несколько недель после инфицирования и сохраняются в крови выздоровевшего до 2 лет.
Подготовка к исследованию
Специальной подготовки не требуется.
Показания к исследованию
Для диагностики заболеваний верхних дыхательных путей хламидийной этиологии.
Для дифференциальной диагностики пневмоний.
Для ретроспективного подтверждения инфекции, вызванной C. pneumoniaе.
Интерпретация
Референсные значения – отрицательно
Положительный результат
Текущее заболевание, вызванное C. pneumoniaе.
Ранее перенесенная инфекция, вызванная C. pneumoniaе.
Отрицательный результат
Отсутствие инфицирования бактерией C. pneumoniaе.
Низкий или неопределяемый уровень антител к C. pneumoniaе в крови:
ранний период заболевания,
длительный срок после инфицирования (более двух лет).
На результаты могут влиять
На результаты анализа способны повлиять нарушения со стороны иммунной системы, аутоиммунные заболевания, ВИЧ.
Назначается в комплексе с
Антитела к Chlamydia pneumoniae, Ig M
Chlamydia pneumoniae, ДНК, ПЦР в плазме крови
Chlamydia pneumoniae – Chlamydia pneumoniae
Виды бактерий
Chlamydia pneumoniae – это разновидность Chlamydia , облигатной внутриклеточной бактерии, которая поражает людей и является основной причиной пневмонии . Он был известен как тайваньский острый респираторный агент (TWAR) по названиям двух исходных изолятов – Тайвань (TW-183) и изолята острого респираторного заболевания, обозначенного AR-39. Вкратце, он был известен как Chlamydophila pneumoniae, и в некоторых источниках это название используется как альтернативное. В некоторых случаях во избежание путаницы даются оба имени.
C. pneumoniae имеет сложный жизненный цикл и для воспроизводства должна инфицировать другую клетку ; таким образом, он классифицируется как облигатный внутриклеточный патоген . Полная последовательность генома C. pneumoniae была опубликована в 1999 году. Он также заражает и вызывает болезни коал , удавов изумрудного дерева ( Corallus caninus ), игуан , хамелеонов , лягушек и черепах.
Первым известным случаем заражения C. pneumoniae был конъюнктивит на Тайване в 1950 году. В истории человечества до 1950 года не было известных случаев C. pneumoniae . Эта атипичная бактерия обычно вызывает фарингит , бронхит , ишемическую болезнь сердца и атипичную пневмонию. в дополнение к нескольким другим возможным заболеваниям.
Микрофотография Chlamydia pneumoniae в эпителиальной клетке при остром бронхите: 1 – инфицированный эпителиоцит, 2 – неинфицированные эпителиоциты, 3 – хламидийные тельца в клетке, 4 – ядра клеток.Жизненный цикл и способ заражения
Жизненный цикл Chlamydia pneumoniae . А – Элементарное тело хламидии . Б – Клетка легкого. 2 – В клетку попадает хламидиоз . 3 – Элементарное тело становится сетчатым телом. 4 – Репликация. 5 – Сетчатые тела становятся элементарными телами и высвобождаются для заражения других клеток.Chlamydia pneumoniae – это небольшая грамотрицательная бактерия (от 0,2 до 1 мкм ), которая претерпевает несколько трансформаций в течение своего жизненного цикла. Он существует как элементарное тело (EB) между хостами . EB не является биологически активным, но устойчив к стрессам окружающей среды и может выживать вне хозяина в течение ограниченного времени. БЭ переносится от инфицированного человека в легкие неинфицированного человека небольшими каплями и несет ответственность за инфекцию. Попадая в легкие, EB поглощается клетками в мешочке, называемом эндосомой , в результате процесса, называемого фагоцитозом . Однако EB не разрушается при слиянии с лизосомами , как это типично для фагоцитированного материала. Вместо этого он трансформируется в сетчатое тело (RB) и начинает реплицироваться внутри эндосомы. Сетчатые тела должны использовать часть клеточного метаболизма хозяина для завершения своей репликации. Затем сетчатые тела превращаются обратно в элементарные тела и попадают обратно в легкие, часто после смерти клетки-хозяина. После этого EB могут инфицировать новые клетки либо в том же организме, либо в новом хозяине. Таким образом, жизненный цикл C. pneumoniae делится между элементарным телом, которое способно инфицировать новых хозяев, но не может реплицироваться, и сетчатым телом, которое реплицируется, но не способно вызывать новую инфекцию.
Болезни
C. pneumoniae – частая причина пневмонии во всем мире; он обычно передается здоровым людям и является формой внебольничной пневмонии . Его лечение и диагностика отличаются от исторически признанных причин, таких как Streptococcus pneumoniae . Поскольку он плохо окрашивает грамм, и поскольку бактерии C. pneumoniae сильно отличаются от многих других бактерий, вызывающих пневмонию (раньше это даже считалось вирусом), пневмония, вызванная C. pneumoniae , классифицируется как « атипичная пневмония ».
Один мета-анализ из серологических данных , сравнивающих перед С. пневмонии инфекции у пациентов с и без рака легких найдены результаты предполагающие перед инфекцией было связано с повышенным риском развития рака легких.
В исследовании , в связь между C. пневмонией инфекцией и атеросклерозом и ишемической болезнью сердца , серологическим тестированием, прямым гистологическим анализом бляшек, и в пробирке тестировании предполагает инфицирование C. пневмонии является существенным фактором риска развития атеросклеротических бляшек и атеросклероза. Инфекция C. pneumoniae увеличивает прилипание макрофагов к эндотелиальным клеткам in vitro и аорте ex vivo . Однако большинство текущих исследований и данных недостаточны и не определяют, как часто C. pneumoniae обнаруживается в атеросклеротической или нормальной ткани сосудов .
C. pneumoniae также была обнаружена в спинномозговой жидкости пациентов с диагнозом рассеянный склероз.
Инфекция C. pneumoniae была впервые связана с хрипом, астматическим бронхитом и астмой у взрослых в 1991 году. Последующие исследования жидкости бронхоальвеолярного лаважа у педиатрических пациентов с астмой, а также с другими тяжелыми хроническими респираторными заболеваниями показали, что более 50 процентов имели доказательства C. pneumoniae путем прямой идентификации микроорганизмов. Инфекция C. pneumoniae вызывает острое дыхание, если оно становится хроническим, диагностируется астма. Эти наблюдения предполагают, что острая инфекция C. pneumoniae способна вызывать разнообразные проявления хронического респираторного заболевания, ведущего к астме.
Инфекция C. pneumoniae также связана с шизофренией. Многие другие патогены связаны с шизофренией.
Лечение макролидными антибиотиками может улучшить состояние при астме в подгруппе пациентов, которую еще предстоит четко определить. Преимущества макролидов впервые были предложены в двух наблюдательных и двух рандомизированных контролируемых исследованиях лечения астмы азитромицином. Одно из этих РКИ и другое исследование макролидов предполагают, что эффект лечения может быть наибольшим у пациентов с тяжелой рефрактерной астмой. Эти клинические результаты коррелируют с эпидемиологическими данными о том, что C. pneumoniae положительно связана с тяжестью астмы, и лабораторными доказательствами того, что инфекция C. pneumoniae создает резистентность к стероидам. Недавний мета-анализ 12 РКИ макролидов для длительного лечения астмы обнаружил значительное влияние на симптомы астмы, качество жизни, гиперреактивность бронхов и пиковую скорость потока, но не на ОФВ1. Данные рандомизированных контролируемых исследований макролидов пациентов с неконтролируемой тяжелой и рефрактерной астмой будут иметь решающее значение для определения роли макролидов при астме.
Исследования вакцин
В настоящее время нет вакцины для защиты от Chlamydia pneumoniae . Идентификация иммуногенных антигенов имеет решающее значение для создания эффективной субъединичной вакцины против инфекций C. pneumoniae . Кроме того, во всем мире наблюдается общая нехватка средств для выявления / диагностики Chlamydia pneumoniae .
Рекомендации
Внешние ссылки
Хламидиоз, симптомы и лечение хламидиоза в клинике ИнТайм
Хламидиоз – опасное заболевание, вызванное возбудителем Chlamydia trachomatis, которым легко заразиться, но сложно диагностировать.
Сложность диагностики заключается в том, что хламидиоз не обладает ярко выраженными характерными именно ему симптомами, картина заболевания может быть схожа со многими урогенитальными инфекциями. Хламидиоз у мужчин может вообще не проявляться никакими симптомами, однако при этом заболевание остается одним из наиболее опасных и ведущих ко многим осложнениям мочеполовой сферы.
Квалифицированные специалисты клиники «ИнТайм» готовы провести качественную диагностику, позволяющую со 100% точность выявить в организме бактерии Chlamydia trachomatis. В нашей клинике применяются и дают высокие результаты наиболее прогрессивные схемы лечения хламидиоза, используемые во всем мире.
Регулярные осмотры врачей-гинекологов и урологов медицинского центра «ИнТайм» — это гарантия своевременного выявления заболеваний, передающихся половым путем, и эффективное и быстрое избавление от болезни.
Почему стоит выбрать клинику «ИнТайм» для лечения хламидиоза?
Опытные врачи
В клинике «ИнТайм» ведут прием врачи высшей категории, гинекологи и урологи с большим опытом успешного лечения хламидиоза у мужчин и женщин. При необходимости можно получить консультацию профессора или академика медицинских наук, которые также работают в клинике «ИнТайм».
Собственная лаборатория
В условиях современной клинико-диагностической лаборатории имеется возможность выполнять самые сложные виды исследований. Для выявления урогенитального хламидиоза применяют метод ПЦР диагностики. Многие результаты могут быть готовы в течение 10-15 минут, однако для некоторых исследований требуется до 3 дней.
Комфорт и удобство
В клинике создана дружественная атмосфера и комфортные условия для пребывания пациентов. Попасть на прием можно в любое удобное время по предварительной записи. Удобное расположение клиники недалеко от станций метро Парк Культуры, Фрунзенская и Спортивная позволяет добраться к нам любым видом транспорта.
Отправьте заявкуна бесплатную консультацию
Мы осуществляем первую бесплатную консультацию для новых пациентов. Просто оставьте свои контактные данные, и мы свяжемся с вами в течение рабочего дня.
Если вы уже были нашим клиентом, вы можете воспользоваться этой формой для заказа обратного звонка.
Спасибо!
Ваша заявка принята.
Мы свяжемся с вами в течение рабочего дня.
Как лечить хламидиоз?
Инфекционное заболевание хламидиоз – является одним из наиболее распространенных заболеваний, которое передается половым путем. Часто течение хламидиоза у женщин не сопровождается никакими симптомами, иногда ощущается чувство жжения и зуд в области внешних и внутренних половых органов, могут появиться боли внизу живота.
Заражение хламидиями происходит во время незащищенного полового акта, также ребенок может инфицироваться данным видом бактерий при прохождении через родовые пути, если у мамы был выявлен хламидиоз при беременности.
После выявления и лабораторного подтверждения диагноза хламидиоз, необходимо обоим партнерам воздержаться от половых отношений и строго соблюдать назначенную врачом схему лечения хламидиоза для женщины и мужчины. Лечение состоит из курса антибактериальных средств, витаминных комплексов, препаратов, стимулирующих выработку интерферона и оказывающих заживляющее действие на ткани.
«ИнТайм» – гарантия точного диагноза и эффективное лечение хламидиоза
Длительное игнорирование хламидиоза может привести к серьезным патологиям женской и мужской половой сферы. Врачи клиники «ИнТайм» рекомендуют проводить профилактический осмотр и ПЦР диагностику на выявления хламидий как минимум раз в год, а при частой смене партнера – не реже 2-х раз в год.
Обращайтесь в медицинский центр «ИнТайм», здесь вам окажут квалифицированную медицинскую помощь и своевременно выявят возможных возбудителей заболеваний.
| |||||
| Chlamydophila pneumoniae (Grayston et al. 1989) Everett et al. 1999 | |||||
| |||||
Chlamydia Pneumoniae Infections – HealthyChildren.org
Когда вы слышите слово chlamydia, вы можете подумать о заболевании, передающемся половым путем (ЗППП) под этим названием. ЗППП вызывается Chlamydia trachomatis, одним видом из бактерий Chlamydia . Другой вид, называемый Chlamydia (или Chlamydophila ), pneumoniae, , вызывает респираторные заболевания. Эти легочные инфекции распространяются так же, как и многие другие респираторные заболевания.Они передаются от человека к человеку напрямую через кашель или чихание и косвенно через микробы на руках или других предметах. Пик числа этих инфекций приходится на детей школьного возраста от 5 до 15 лет.
Признаки и симптомы
Болезни, вызываемые C pneumoniae , могут вызывать продолжительный кашель, бронхит и пневмонию, а также боль в горле, ларингит, ушные инфекции и синусит. Обычно они начинаются постепенно с боли в горле, которая сменяется кашлем примерно через неделю или более.Кашель может длиться от 2 до 6 недель. В некоторых случаях ребенок может заболеть бронхитом или легкой формой пневмонии. В то время как у некоторых инфицированных детей симптомы проявляются от слабых до умеренных или вообще отсутствуют, у других инфекция может протекать тяжелее.
Как ставится диагноз?
Многие случаи C pneumoniae диагностируются педиатром после физического осмотра ребенка и изучения его симптомов. Врач также может назначить анализы крови, которые обнаруживают антитела к бактериям.Однако может потребоваться неделя или больше, чтобы антитела проявились в крови. Хотя существуют специальные лаборатории, которые могут оценивать образцы мазков из носа или горла, в настоящее время нет надежных коммерчески доступных исследований.
Лечение
Выздоровление от респираторной инфекции Chlamydia может быть медленным. Ваш педиатр может назначить антибиотики, такие как эритромицин или тетрациклин, чтобы избавиться от инфекции и помочь вашему ребенку быстрее поправиться.
Профилактика
Чтобы снизить вероятность заражения вашего ребенка инфекцией C pneumoniae , ему следует соблюдать правила гигиены, включая частое мытье рук.
Информация, содержащаяся на этом веб-сайте, не должна использоваться вместо медицинской помощи и рекомендаций вашего педиатра. Ваш педиатр может порекомендовать лечение по-разному, исходя из индивидуальных фактов и обстоятельств.
Chlamydia Pneumoniae (Chlamydophila) – Консультант по инфекционным заболеваниям
ОБЗОР: Что нужно знать каждому врачу
Название и классификация возбудителя
Chlamydia pneumoniae – внутриклеточный организм с клеточной мембраной, но без клеточной стенки
Во многих исследованиях это третья или четвертая по частоте причина внебольничной пневмонии (ВП).Однако, учитывая трудности в постановке точного этиологического диагноза инфекции, вызванной этим микроорганизмом, все еще существуют некоторые разногласия относительно того, является ли это основным патогеном при ВП.
Часто встречаются сочетанные инфекции S. pneumoniae и M. pneumoniae.
Продолжить чтение
C. pnemoniae вызывает целый ряд хронических заболеваний, таких как атеросклероз, астма, артрит, рассеянный склероз и многие другие. Однако доказательства наличия причинно-следственной связи в большинстве из них неубедительны.
Большинство эмпирических методов лечения ВП антибиотиками относятся к C. pneumoniae.
Хламидии – облигатные внутриклеточные бактериальные патогены. Недавний таксономический анализ рода Chlamydia показал, что существует два рода – Chlamydia и Chlamydophila. Хламидофила имеет следующие виды: C. пекорум, вызывающий инфекцию у крупного рогатого скота, овец и коал; C. pneumoniae и C. psittaci; C. abortus, вызывающий аборт у овец и крупного рогатого скота; C. cavieae, вызывающая конъюнктивит морских свинок; и С.felis, вызывающий кератоконъюнктивит у кошек.
C. pneumoniae имеет грамотрицательную клеточную стенку и уникальный цикл развития с элементарными (инфекционная частица) и сетчатыми (внутриклеточная репликативная частица) телами. После заражения элементарные тельца (диаметром 200-400 нм) прикрепляются к клетке-хозяину за счет электростатического связывания и проникают в клетку посредством эндоцитоза. Элементарные тела (EB) дифференцируются в сетчатые тела (RB), которые подвергаются двойному делению. Примерно через 36 часов сетчатые тела (RB) снова дифференцируются в элементарные тела (EB).В клетке накапливается большое количество EB (примерно 500-1000), но функция клетки-хозяина не нарушается. Эти большие скопления или микроколонии называют внутрицитоплазматическими включениями. Во время этого процесса хламидийные антигены высвобождаются на поверхность клетки-хозяина, вызывая иммунный ответ хозяина. Высвобождение ЭБ происходит с помощью нескольких процессов, включая экструзию включения. Также вероятно, что C. pneumoniae переходит в устойчивое состояние, когда он метаболически неактивен и на этой стадии не подвержен влиянию антибактериальной терапии.Гамма-интерферон, антибиотики и недостаток питательных веществ являются вероятными факторами персистенции C. pneumoniae.
C. pneumoniae была обнаружена в ходе исследований трахомы в 1960-х годах доктором Т. Грейстоном и его коллегами. Первоначально эти исследователи назвали недавно обнаруженный организм TWAR в честь своих первых двух изолятов TW-183 (TW для Twain) и AR-39 (AR для острых респираторных заболеваний). Существует только один серотип C. pneumonia, и все изоляты от людей на сегодняшний день имеют высокий процент (> 98%) родства.
C. pneumoniae – единственный вид, у которого есть ген udk, кодирующий уридинкиназу. Секвенирование штамма коалы C.pneumoniae позволяет предположить, что люди заразились этой инфекцией зоонозно.
Какое лечение лучше всего?
Предпочтительные противомикробные агенты включают Доксициклин 100 два раза в день в течение 14-21 дней; азитромицин 1,5 г перорально в течение 5 дней; кларитромицин 500 мг 2 раза в день в течение 10 дней; левофлоксацин 500 мг перорально или внутривенно 1 раз в сутки в течение 7-14 дней.
Данные по лечению ограничены.Исследования с использованием культуры показали, что макролиды (эритромицин, кларитромицин, азитромицин) и хинолоны (левофлоксацин и моксифлоксацин) уничтожали микроорганизмы в 70-86% случаев. Клиническое улучшение обычно наступает, несмотря на упорство организма.
Устойчивость к хинолонам in vitro связана с точечной мутацией в гене GyrA.
Как пациенты заражаются этой инфекцией и как предотвратить ее распространение среди других пациентов?
Эпидемиология
С.pneumoniae – распространенный респираторный патоген. Инфекция часто происходит в детстве и подростковом возрасте, а к зрелому возрасту инфицировано около 80% населения. Бессимптомные респираторные инфекции встречаются у 2-5% взрослых и детей. Роль такой инфекции неясна. Нет сезонных колебаний, хотя есть предположение о четырехлетнем цикле заболеваемости C. pneumoniae pneumonia. Пневмония, вызванная этим организмом, встречается во всем мире. В одном исследовании на него приходилось 8% случаев в Северной Америке, 7% в Европе, 6% в Латинской Америке и 5% в Азии / Африке.Ставки наиболее высоки среди лиц моложе 1 года и старше 70 лет.
Часто встречаются сочетанные инфекции S. pneumoniae и M. pneumoniae.
Взрослые, серопозитивные по C. pneumoniae, с большей вероятностью имели более высокий индекс массы тела, курили в настоящее время и имели хроническую обструктивную болезнь легких и более высокое систолическое артериальное давление, чем серонегативные субъекты.
Заражение через капельное заражение произошло во время лабораторной аварии.C. pneumoniae может выжить на столешнице до 30 часов. Описано распространение в семьях, призывниках и домах престарелых. Интересно, что по крайней мере в одной вспышке в доме престарелых те, кто имел более высокий уровень физической активности, были более подвержены заражению. Контакт с инфицированными обитателями дома престарелых был фактором риска заражения персонала.
Нет сезонных различий в заболеваемости C. pneumonia, и заболеваемость остается стабильной.
Проблемы инфекционного контроля
В повседневной практике нет проблем с инфекционным контролем, поскольку редко кто знает, что инфекция вызвана этим микроорганизмом.Однако в случае вспышки, вызванной C. pneumoniae, или если вы знаете, что респираторная инфекция вызвана этим организмом, следует соблюдать респираторные меры предосторожности.
В настоящее время вакцины нет, и противоинфекционная профилактика не рекомендуется.
Каковы клинические проявления заражения этим организмом?
Наиболее частыми проявлениями являются инфекции дыхательных путей, проявляющиеся в виде пневмонии или фарингита.
Инкубационный период 21 день.Продолжительность симптомов до госпитализации составляла 4,6 дня для C. pneumoniae по сравнению с 12,9 дней для пациентов с другими причинами пневмонии. Заболевание варьируется от легкой до очень тяжелой. Большинство случаев имеют степень тяжести от легкой до средней степени тяжести. Симптомы неспецифичны. Охриплость голоса чаще связана с пневмонией, вызванной C. pneumoniae, чем с другими респираторными патогенами. Лихорадка может сопровождаться миалгией и ознобом, а затем кашлем. Кашель, вероятно, является наиболее заметным признаком болезни. Кашель и недомогание могут сохраняться в течение нескольких месяцев, несмотря на терапию антибиотиками.Это может вызвать пневмонию у хозяина с ослабленным иммунитетом.
При сравнении симптомов и признаков 18 пациентов с серологическим диагнозом C. pneumoniae с 282 пациентами с внебольничной пневмонией, у которых были отрицательные антитела к C. pneumoniae, не было выявлено различий в симптомах или клинических проявлениях. В этом исследовании 61% пациентов с C. pneumoniae имели количество лейкоцитов более 10 000 на микролитр. Во многих описаниях пневмонии, вызванной C. pneumoniae, количество лейкоцитов считается нормальным.Продолжительность пребывания в больнице для пациентов с C. pneumoniae была больше – 9 плюс-минус 7,3 дня по сравнению с 7,58 плюс-минус 3,49 дня для пациентов с C. pneumoniae и сопутствующим патогеном. Смертность от C. pneumoniae составила 4,9%, что незначительно отличалось от остальных случаев пневмонии.
В одном исследовании у пациентов с пневмонией, вызванной C. pneumoniae, застойная сердечная недостаточность была выше, чем у пациентов с пневмококковой пневмонией. Это может быть отражением более старшего возраста, поскольку средний возраст C.pneumoniae и S.pneumoniae соответственно.
Рентгенологические проявления C. pneumoniae во многом такие же, как показано выше, и не являются специфическими для этого заболевания. При обзоре компьютерно-томографических характеристик 40 пациентов с пневмонией, вызванной C. pneumoniae, по сравнению с 42 пациентами с пневмонией Mycoplasma pneumoniae, были выявлены следующие особенности:
Затухание в матовом стекле (n = 38) и ацинарные паттерны (n = 28) были основными результатами компьютерной томографии при пневмонии C. pneumoniae.
Ацинарный паттерн и плевральный выпот чаще наблюдались у пациентов с пневмонией, вызванной M. pneumoniae.
Центрилобулярные узелки и утолщение бронхиальной стенки встречались реже, чем при пневмонии M. pneumoniae.
C. pneumoniae и хронические болезни
Астма
Хроническая стойкая инфекция C. pneumoniae связана с астмой, артритом и атеросклерозом.
В начале 1990-х рабочие в США сообщили о связи между серологическими признаками острой C.pneumoniae и астма. С тех пор в разных странах был проведен ряд исследований, чтобы определить, является ли это истинной ассоциацией. Интраназальная инокуляция мышей C. pneumoniae приводит к стойкой гиперчувствительности дыхательных путей и воспалению дыхательных путей. В недавнем систематическом обзоре делается вывод, что C. pneumoniae, по-видимому, больше связана с персистированием астмы, чем с обострениями. Кокрановский обзор лечения хронической астмой марколидом выявил общий положительный эффект на симптомы и эозинофильные маркеры воспаления.
Атеросклероз
Воспаление – важный, но малоизученный фактор развития атеросклероза. Традиционные факторы риска гипертонии, дислипидемии, табакокурения, ожирения и семейного анамнеза не полностью объясняют, как воспаление способствует прогрессированию атеросклероза. Таким образом, внимание обратилось на возможность того, что микробные инфекции и иммунологические механизмы играют роль в патогенезе атеросклероза.Исследование случай-контроль, проведенное в Финляндии в 1988 году, показало, что пациенты с ишемической болезнью сердца чаще имеют антитела к C. pneumoniae, чем контрольные субъекты. С тех пор было проведено множество исследований, пытающихся установить, что C. pneumoniae действительно играет роль в развитии атеросклероза. Серологические исследования и исследования изоляции не подтвердили причинно-следственную связь. В значительной степени это связано с межлабораторными различиями в используемых методах. Например, загрязнение часто встречается при вложенной полимеразной цепной реакции (ПЦР), но в меньшей степени при ПЦР в реальном времени.В более позднем подходе использовался количественный анализ липополисахарида хламидии. При использовании этого подхода была выявлена корреляция между уровнями cLPS в сыворотке крови и уровнями С-реактивного белка при остром инфаркте миокарда и нестабильной стенокардии.
Другой подход – использовать терапию антибиотиками, чтобы повлиять на исход ишемической болезни сердца. Мета-анализ 11 рандомизированных испытаний, в которых приняли участие 19 217 пациентов, которые использовали различные антибиотики и разную продолжительность наблюдения, не выявил эффекта от антибиотиков, более того, пять из более крупных исследований отдают предпочтение плацебо.
Исследователи сейчас используют протеомный подход к этой проблеме. Восемь (RpoA, MOMP, YscC, Pmp10, PorB, Pmp21-m, GroEL и Cpaf-c) из 31 антигенов C. pneumoniae показали более высокую реактивность с сыворотками от ПЦР-положительных доноров с использованием метода двумерного гель-иммуноблоттинга. Кроме того, ответ антител у субъектов с положительной реакцией на ДНК C. pneumonia показал реактивность по отношению к антигенам, селективно повышенную во время персистенции C. pneumoniae.
С.pneumoniae и рассеянный склероз
Chlamydia pneumoniae – один из длинного списка (более 20) вирусов и бактерий, вовлеченных в этиологию рассеянного склероза за последние 50 лет. Действительно, самые последние споры в этой области заключаются в том, что расширение и стентирование стенозированных шейных вен приводит к заметному улучшению симптомов рассеянного склероза (РС). Гипотеза состоит в том, что C. pneumoniae действует как кофактор, усиливая уже установленный воспалительный и демиелинизирующий процесс при РС.На сегодняшний день нет доказательств роли C. pneumoniae в развитии рассеянного склероза.
C. pneumoniae и болезнь Альцгеймера
В одном исследовании большая часть образцов головного мозга пациентов с болезнью Альцгеймера была положительной на C. pneumonia в ПЦР, в то время как образцы головного мозга, не относящиеся к контрольным субъектам, не страдающим болезнью Альцгеймера, не были. В одном исследовании пациентам с AD назначался 3-месячный курс доксициклина, и через 6 месяцев в группе лечения наблюдалось меньшее снижение когнитивных функций, чем в контрольной группе.
C. pneumoniae и реактивный артрит
В 1916 году был отмечен воспалительный артрит после эпизода дизентерии. С тех пор несколько видов бактерий, включая C. pneumonia, были вовлечены в реактивный артрит (синдром Рейтера). Тринадцать процентов из 200 пациентов с различными артритами и синовиальной биопсией дали положительный результат на C. pneumoniae с помощью ПЦР. Как и в случае других хронических болезненных состояний, роль C. pneumoniae в реактивном артрите далеко не ясна.
Какие общие осложнения связаны с заражением этим возбудителем?
Как мне идентифицировать организм?
C. pneumoniae имеет уникальный цикл развития с элементарными (инфекционная частица) и сетчатыми (внутриклеточная репликативная частица) телами. После заражения элементарные тельца (диаметром 200-400 нм) прикрепляются к клетке-хозяину за счет электростатического связывания и проникают в клетку посредством эндоцитоза. Элементарные тела дифференцируются в сетчатые тела, которые подвергаются двойному делению.Примерно через 36 часов RB снова дифференцируются в EB. В клетке накапливается большое количество EB (около 500-1000), но функция клетки-хозяина не нарушается. Эти большие скопления или микроколонии называются внутрицитоплазматическими включениями. Во время этого процесса хламидийные антигены высвобождаются на поверхность клетки-хозяина, вызывая иммунный ответ хозяина. Высвобождение ЭБ происходит с помощью нескольких процессов, включая экструзию включения. Также вероятно, что C. pneumoniae переходит в устойчивое состояние, когда он метаболически неактивен и на этой стадии не подвержен влиянию антибактериальной терапии.Гамма-интерферон, антибиотики и недостаток питательных веществ являются вероятными факторами персистенции C. pneumoniae.
C. pneumoniae можно выделить в культуре ткани с использованием клеток HEp-2. Образцы мазков из носоглотки или глотки, собранные с помощью тампонов из дакрона с алюминиевым стержнем или пластмассовым стержнем, имеют важное значение, потому что альгинат кальция или ватные наконечники и деревянные стержни могут подавлять рост микроорганизмов в культуре ткани и могут быть токсичными для клеток. Через 72 часа инкубации рост можно определить с помощью иммунофлуоресцентного красителя.Для обнаружения роста может потребоваться более одного прохода.
ПЦР все еще является исследовательским тестом.
Тест на микроиммунофлуоресценцию (MIF), вероятно, лучший. Иммуноферментный анализ (ИФА) легче выполнить. Поскольку C. pneumoniae является внутриклеточным организмом, неудивительно, что существует слабая корреляция между прямым обнаружением организма с помощью выделения или ПЦР и серологией. Трудности с использованием серологии для этиологического диагноза инфекции C. pneumoniae иллюстрируются двумя исследованиями пневмонии, в которых у 7-13% пациентов были положительные культуры на C.pneumoniae и 7-18% имели положительную серологию MIF. Проблема только в том, что это были разные пациенты.
Точный лабораторный диагноз инфекции C. pneumoniae зависит от высокой квалификации лабораторных работников и доступа к средствам для культивирования тканей.
Как этот организм вызывает болезнь?
По большей части механизмы того, как C. pneumoniae вызывает повреждение тканей, неизвестны. Однако о том, как C.trachomatis вызывает повреждение тканей, и многие из тех же механизмов могут действовать в отношении C. pneumoniae. Хламидийный белок теплового шока 60 (hsp 60) вызывает антительный ответ, связанный с повреждением глаз и половых путей, когда C. trachomatis заражает эти области. Есть предположение, что продолжающийся синтез hsp 60 во время латентного периода C. pneumoniae в дыхательных путях может быть связан с персистированием астмы.
КАКИЕ ДОКАЗАТЕЛЬСТВА для конкретных рекомендаций по ведению и лечению?
Hammerschlag, MR.«Достижения в борьбе с инфекциями». Expert Rev Anti-Infect Ther. т. 1. 2003. С. 493-504. (Этот источник суммирует данные многоцентровых исследований, в которых использовались культуры для диагностики C. pneumoniae. Лечение эритромицином, кларитромицином, азитромицином, левофлоксацином и моксифлоксацином было примерно на 70-80% эффективным в уничтожении C. pneumoniae из носоглотки у детей и взрослых с внебольничная пневмония.)
Copyright © 2017, 2013 ООО «Поддержка принятия решений в медицине».Все права защищены.
Ни один спонсор или рекламодатель не участвовал, не одобрял и не платил за контент, предоставляемый Decision Support in Medicine LLC. Лицензионный контент является собственностью DSM и защищен авторским правом.
Chlamydia pneumoniae часто обнаруживается в крови после острой легочной инфекции.
В редакцию:
Облигатная внутриклеточная бактерия Chlamydia pneumoniae – частая причина острых респираторных инфекций с распространенностью во всем мире до 70% 1.Было высказано предположение, что персистентность бактерий после острой инфекции в дыхательных путях или в атеросклеротических кровеносных сосудах участвует в патогенезе хронических воспалительных заболеваний, таких как хроническая обструктивная болезнь легких и атеросклероз 2, 3. На животных моделях было показано, что после острого Инфекция легких C. pneumoniae , патоген системно распространяется в кровотоке с использованием моноцитов крови в качестве вектора 4, 5. Однако данные, подтверждающие эту гипотезу у людей, все еще отсутствуют.Из наблюдений in vitro и известно, что инфицирование C. pneumoniae моноцитов крови человека приводит к нерепликативному, но жизнеспособному состоянию патогена, который невосприимчив к лечению антибиотиками 6.
Целью данного исследования был анализ распространяется ли C. pneumoniae системно из легких в кровь у пациентов с острой инфекцией нижних дыхательных путей (LRTI). Пациенты, выздоравливающие после острой инфекции легких, вызванной C. pneumoniae , наблюдались на наличие C.pneumoniae в крови во время контрольных визитов (через 30 и 90 дней после инфицирования) и по сравнению с пациентами после инфицирования Streptococcus pneumoniae .
Пациенты, поступающие в отделение неотложной помощи или амбулаторные отделения университетской больницы Шлезвиг-Гольштейн, кампус Любек (Любек, Германия) с рентгенологически подтвержденной нетяжелой внебольничной пневмонией (ВП) или тяжелой ИДПТ с лихорадкой, продуктивным кашлем и положительными результатами аускультативных исследований. протестировано в течение 3-летнего периода на наличие C.pneumoniae или S. pneumoniae в качестве возбудителей болезни. Критериями включения были способность производить мокроту, выполнять функциональный тест легких и готовность вернуться для последующих посещений после выздоровления от острой инфекции; Исключались пациенты с иммунодефицитом и пациенты из домов престарелых. Первоначально было набрано 42 пациента с подтвержденной инфекцией C. pneumoniae или S. pneumoniae (CAP: n = 40; острый бронхит: n = 2).Чтобы преодолеть различия в частотах инфицирования C. pneumoniae и S. pneumoniae при острых респираторных инфекциях, набор пациента S. pneumoniae проводился после набора пациента C. pneumoniae . Трое пациентов с инфекцией C. pneumoniae были исключены из анализа из-за отзыва согласия или потери наблюдения. Клинические характеристики пациентов были задокументированы на 0 день, 30 день (первый визит) и 90 день (второй визит).Образцы дыхательных путей и крови были взяты при поступлении и при каждом последующем посещении с дополнительными тестами функции легких при первом и втором посещении.
Диагностика инфекции, вызванной S. pneumoniae , проводилась на основе анализа культур респираторных образцов и крови, а также обнаружения пневмококкового антигена в моче (Binax NOW ® ; Inverness Medical International, Cranfield, UK). Образцы спонтанной или индуцированной мокроты (10-минутная ингаляция с 3% раствором NaCl) брали при каждом посещении для обнаружения S.pneumoniae (культура кровяного агара в соответствии с рекомендациями Института клинических и лабораторных стандартов (Уэйн, Пенсильвания, США)) или C. pneumoniae (ПЦР). Выделение моноцитов периферической крови (PBMC) из слепых образцов крови с ЭДТА и обнаружение C. pneumoniae с помощью ПЦР выполняли, как описано ранее 6. Протокол исследования был одобрен этическим комитетом Любекского университета (№: 01-148). ). Все пациенты были проинформированы о цели исследования и дали письменное согласие.Статистический анализ проводился с помощью программного обеспечения SPSS (SPSS Inc., Чикаго, Иллинойс, США) с использованием t-критерия для анализа непарных параметрических параметров и U-критерия Манна – Уитни для анализа непарных непараметрических параметров. Номинальные масштабированные параметры анализировали с использованием критерия хи-квадрат.
Всего в исследование были включены 39 пациентов. У 18 пациентов C. pneumoniae было обнаружено в мокроте как возбудитель острой респираторной инфекции, в том числе 16 пациентов с ВП и 2 пациента с острым бронхитом.21 пациент с инфекциями S. pneumoniae соответствовал критериям ВП. Характеристики пациентов в обеих группах не различались, за исключением того, что пациенты в группе S. pneumoniae были значительно старше (p <0,05) (таблица 1). На момент поступления пациенты, страдающие инфекцией S. pneumoniae , имели сравнительно более высокие системные воспалительные маркеры, например. C-реактивный белок и количество лейкоцитов.
Таблица 1– Характеристики пациента на момент поступленияДля исследования системного распространения C.pneumoniae , образцы крови для C. pneumoniae ПЦР были взяты в течение 30 и 90 дней (посещение один и два, соответственно) после заражения. PBMC пациентов, выздоравливающих от острой инфекции легких C. pneumoniae , были значительно чаще положительными на хламидийную ДНК в течение 30 дней после инфицирования по сравнению с контрольной группой S. pneumoniae (72,7% против 17%, p = 0,007) (рисунок 1). Ранее было показано, что обнаружение ДНК C. pneumoniae в циркулирующих лейкоцитах с помощью ПЦР является ценным инструментом для идентификации C.pneumoniae носителей 7. Общая распространенность C. pneumoniae положительная ДНК была обнаружена у 18,5% здоровых доноров крови и до 59% у пациентов, страдающих ишемической болезнью сердца 8, 9. Наши данные показывают, что частота обнаружения C y. pneumoniae в крови быстро увеличивается (через 30 дней после заражения), но не увеличивается (через 90 дней после заражения) после острой инфекции легких, вызываемой C. pneumoniae (рис. 1).
Рисунок 1–Частота обнаружения ДНК Chlamydia pneumoniae в моноцитах периферической крови (PBMC) пациентов после острой респираторной инфекции C.pneumoniae (▪) или Streptococcus pneumoniae (▴). Фракции PBMC образцов цельной крови были значительно чаще C. pneumoniae ДНК-положительными (ПЦР) через 30 дней после инфицирования (визит 1) у пациентов с острой инфекцией C. pneumoniae по сравнению с пациентами с инфекцией S. pneumoniae . # : p = 0,007.
Инфекции, вызванные C. pneumoniae , составляют 0,9–13% ВП во всем мире и, по всей видимости, появляются эпидемически в течение многих лет 10.В связи с этим проспективные исследования, анализирующие исходы у пациентов, страдающих острой респираторной инфекцией C. pneumoniae с течением времени, трудны для выполнения и до сих пор отсутствуют. Модели на животных предполагают, что C. pneumoniae может распространяться из легких в кровоток после первоначального инфицирования нижних дыхательных путей с использованием моноцитов крови в качестве вектора 4. Поскольку ДНК C. pneumoniae постоянно обнаруживается в различных пропорциях у бессимптомных здоровых людей. Пациенты, клиническая значимость персистенции хламидий в легких, а также в мононуклеарных клетках крови, полностью не изучена 11.Таким образом, C. pneumoniae может сохраняться в легких в течение многих месяцев после острой инфекции, несмотря на длительные курсы терапии тетрациклином или доксициклином 10. Напротив, Blasi et al. 12 показали, что у пациентов с хроническим бронхитом легкой и средней степени тяжести обнаружение ДНК C. pneumoniae в PBMC было связано с более высокой частотой обострений. Они также показали, что устойчивый клиренс C. pneumoniae из крови после лечения азитромицином был достигнут только у 29% пациентов 12.Это может быть связано с низкой восприимчивостью стойких хламидий к терапии макролидами 6, но также может указывать на системное распространение мертвых C. pneumoniae внутри PBMC из хронического резервуара, поскольку внутриклеточный рост и потомство C. pneumoniae серьезно нарушено. в первичных моноцитах и макрофагах, происходящих из моноцитов 13.
Ограниченный размер нашего исследования не позволяет сделать однозначных выводов относительно взаимосвязи между диссеминацией C. pneumoniae и клиническим течением.Однако выздоровление от пневмонии и изменение лечения не различались между пациентами с обнаружением ДНК C. pneumoniae в крови или без нее. Кроме того, между обеими группами не наблюдалось значительных изменений функции легких или высвобождения провоспалительных цитокинов в мокроте или крови с течением времени (данные не показаны).
В заключение, системное распространение C. pneumoniae из легких в кровь после острой респираторной инфекции наблюдается у 70% пациентов в течение 30 дней после инфицирования.Однако клиническая значимость этих результатов в отношении разрешения болезни и патогенеза хронических сосудистых и легочных заболеваний требует дальнейших исследований.
Благодарности
Мы благодарны У. Кнупперцу и Х. Рихарцу (Медицинская клиника III, Университетская клиника Шлезвиг-Гольштейн, кампус Любек, Германия) за отличную техническую поддержку.
Сноски
Заявление о поддержке
Эта работа была поддержана Федеральным министерством образования и исследований Германии (Bundesministerium für Bildung und Forschung), Competence Network CAPNETZ (TP A6) и Европейской инициативой по борьбе с хламидийными инфекциями с помощью объективной геномики (ECIBUG) в рамках ERANET PathoGenoMics (BMBF).
Заявление о заинтересованности
Не объявлено.
Chlamydophila pneumoniae – обзор
Chlamydophila pneumoniae
В последние годы Chlamydophila pneumoniae признано все более частым респираторным патогеном как у детей, так и у взрослых. Ранее известный как агент тайваньского острого респираторного агента (TWAR) и помещенный в тот же род, что и Chlamydia trachomatis, , теперь он помещен в отдельный род, Chlamydophila, вместе с Cp.psittaci . В результате усовершенствованных методов диагностики и повышения осведомленности медицинских работников он получил признание в качестве отдельного патогена и был причастен ко многим острым и хроническим состояниям.
Microbiology
Chlamydophila pneumoniae является генетически, морфологически и антигенно отличным членом семейства Chlamydiaceae. Хламидии – облигатные внутриклеточные патогены с клеточной стенкой, содержащей липополисахариды, похожие на грамотрицательные бактерии. 28 , 29 Хламидии проявляют две формы в течение своего цикла развития: внутриклеточную делящуюся форму (сетчатое тело) и внеклеточную, инфекционную, метаболически неактивную форму (элементарное тело). 28 Ср. pneumoniae демонстрирует уникальное элементарное тело грушевидной формы, которое отличает его от других Chlamydiae . 29
Эпидемиология
Chlamydophila pneumoniae инфекции являются распространенными.Пик первоначальной инфекции наблюдается у детей в возрасте от 5 до 15 лет в развитых регионах с умеренным климатом, но может возникать раньше у детей, проживающих в тропических или менее развитых регионах. Доказательств сезонности нет, хотя эпидемии случаются каждые 4–5 лет. Инфекция, вероятно, передается через респираторные выделения от человека к человеку или через фомиты. 30 Не существует резервуара для животных, хотя организм был изолирован от нескольких видов. По оценкам, от 2% до 5% взрослых и детей являются бессимптомными носоглоточными носителями Cp.pneumoniae , но неясно, являются ли носители резервуаром инфекции. Сопутствующее инфицирование M. pneumoniae или S. pneumoniae встречается часто. Инкубационный период – 21 день. 31
Патология
После доставки на слизистую оболочку дыхательных путей Cp. pneumoniae переносятся в эпителиальные клетки слизистой оболочки путем эндоцитоза. Хламидии предотвращают слияние фагосомы с лизосомами, тем самым подавляя их собственную смерть.Внутри фагосомы элементарные тела дифференцируются в сетчатые тела и подвергаются бинарному делению. 28 Через 36–48 часов после инокуляции размножение прекращается, и сетчатые тела возвращаются в инфекционные элементарные тела. Содержимое фагосомы высвобождается путем цитолиза, экзоцитоза или экструзии всего включения. 28
Клинические проявления
Chlamydophila pneumoniae Инфекции часто начинаются с неспецифического продрома боли в горле, недомогания, головной боли, субфебрильной температуры и кашля.Курс длительный (от 2 до 6 недель) и часто двухфазный. Пациенты могут демонстрировать инфекции верхних дыхательных путей, такие как фарингит, ларингит или синусит. У значительного числа разовьется пневмония или бронхит. Cp. pneumoniae является мощным триггером воспаления и приводит к опосредованной иммуноглобулином E (IgE) бронхиальной реактивности. 32 Физический осмотр может выявить неэкссудативный фарингит, хрипы и мелкие или грубые потрескивания.
Возраст пациента часто определяет степень тяжести заболевания; у подростков чаще развиваются инфекции нижних дыхательных путей. 33 Тяжелое заболевание может поражать ранее здоровых детей и приводить к таким осложнениям, как пневматоцеле, плевральный выпот, пневмоторакс, интерстициальный фиброз или абсцесс легкого. Другие осложнения Cp. pneumoniae включают узловатую эритему, реактивный артрит, синдром Гийена-Барре, менингоэнцефалит, миокардит и эндокардит. Cp. pneumoniae также была выделена из жидкости среднего уха у детей с острым средним отитом, как правило, с другими патогенами.Этот организм был связан с астмой, рассеянным склерозом, хронической усталостью и атеросклеротическим заболеванием сердца; причинно-следственная связь не подтверждена. Смертность часто связана с сопутствующими заболеваниями или вторичными инфекциями.
Лабораторные данные и диагностика
Хотя довольно повсеместно, Cp. pneumoniae диагностируется нечасто из-за неспецифических рентгенологических и лабораторных данных, а также из-за отсутствия надежных диагностических тестов. Количество лейкоцитов часто нормальное или слегка повышенное.Скорость оседания эритроцитов обычно повышена. Рентгенография грудной клетки классически демонстрирует картину альвеолярного инфильтрата или субсегментарного пневмонита без консолидации, 31 , 33-35 , хотя одиночное субсегментарное поражение только одной доли является наиболее распространенным поражением. 31 , 35 Плевральный выпот встречается редко.
Серологическое тестирование является основным методом диагностики. Обнаружение антител с помощью микроиммунофлуоресценции – единственный в настоящее время приемлемый тест для диагностики острой инфекции. 36 , 37 Новые критерии CDC для серологической диагностики требуют титра IgM более 1:16 или четырехкратного повышения титра IgG. 37 Как заражение Cp. pneumoniae может привести только к частичному иммунитету, часто возникают повторные инфекции. Антитела IgM могут отсутствовать при заболевании, связанном с повторной инфекцией, и могут появиться через 3 недели после заболевания, связанного с первичной инфекцией. Не рекомендуется полагаться на один тест для диагностики. 36 , 37 Chlamydophila pneumoniae можно культивировать из носоглотки, мокроты или плевральной жидкости.Образцы необходимо поместить в транспортную среду, кратковременно хранить при 4 ° C и засеять на среды, содержащие клетки. Идентификация бактерий по окрашиванию затруднена, так как Cp. pneumoniae – специальный реагент ограничен. Методы полимеразной цепной реакции не стандартизированы и не одобрены Управлением по контролю за продуктами и лекарствами США (FDA) для коммерческого использования. 37 , 38
Лечение
В нескольких исследованиях изучали реакцию Cp.pneumoniae на противомикробную терапию. Cp. pneumoniae чувствительна к макролидам, тетрациклинам и хинолонам; однако последние два обычно не рекомендуются детям. Детей младшего возраста можно лечить 14-дневным курсом перорального эритромицина, 10-дневным курсом кларитромицина или 5-дневным курсом перорального азитромицина. 36 , 38 Пациентов-подростков можно лечить 5-дневным курсом перорального азитромицина, 14-21-дневным курсом перорального доксициклина или 14-21-дневным курсом перорального тетрациклина. 36 , 38 Может потребоваться длительная противомикробная терапия (до 3 недель). 36 Пациентам с тяжелым заболеванием может потребоваться парентеральная терапия.
Профилактика
Передача Chlamydophila может быть уменьшена путем избегания контакта с людьми с респираторными заболеваниями и их респираторными выделениями. Антибиотики не рекомендуются ни для устранения носительства, ни для постконтактной профилактики. Вакцины против Cp не существует.pneumoniae в это время.
Хроническая инфекция, вызванная Chlamydia pneumoniae при астме: фенотип, связанный с низкой инфекцией, тип 2 | Респираторные исследования
Huang YJ, Boushey HA. Микробиом при астме. J Allergy Clin Immunol. 2015; 135 (1): 25–30.
PubMed PubMed Central Статья Google Scholar
Хан DL. Инфекционная астма: возрождение клинической картины? J Fam Pract. 1995. 41 (2): 153–7.
CAS PubMed PubMed Central Google Scholar
Rantala A, Jaakkola JJK, Jaakkola MS. Респираторные инфекции предшествуют развитию астмы у взрослых. PLoS ONE. 2011; 6 (12): e27912.
CAS PubMed PubMed Central Статья Google Scholar
Webley WC, Хан DL. Инфекционно-опосредованная астма: этиология, механизмы и варианты лечения, с особым вниманием к Chlamydia pneumoniae и макролидам.Respir Res. 2017; 18 (1): 98.
PubMed PubMed Central Статья CAS Google Scholar
Yeh JJ, Wang YC, Hsu WH, Kao CH. Заболевание астмой и Mycoplasma pneumoniae : общенациональное когортное исследование. J Allergy Clin Immunol. 2016; 137 (4): 1017–23.
PubMed Статья PubMed Central Google Scholar
Hahn DL, Anttila T, Saikku P.Ассоциация антител IgA к Chlamydia pneumoniae с недавно появившейся симптоматической астмой. Epidemiol Infect. 1996. 117 (3): 513–7.
CAS PubMed PubMed Central Статья Google Scholar
Blasi F, Allegra L, Tarsia P, Hahn DL. Chlamydia pneumoniae и астма. Грудная клетка. 1998; 53 (12): 1095.
CAS PubMed Статья PubMed Central Google Scholar
Хан Д.Л., Додж Р.В., Голубятников Р. Ассоциация инфекции Chlamydia pneumoniae (штамм TWAR) с хрипом, астматическим бронхитом и астмой у взрослых. JAMA J Am Med Assoc. 1991. 266 (2): 225–30.
CAS Статья Google Scholar
Patel KK, Webley WC. Доказательства инфекционного фенотипа астмы: аллергия, вызванная хламидиозом, и патоген-специфический IgE в модели новорожденных мышей. PLoS ONE. 2013; 8 (12): e83453.
PubMed PubMed Central Статья CAS Google Scholar
Хан DL, Plane MB, Mahdi OS, Byrne GI. Вторичные результаты пилотного рандомизированного исследования лечения астмы азитромицином. Клинические испытания PLoS. 2006; 1 (2): e11.
PubMed PubMed Central Статья CAS Google Scholar
Hahn DL, Пилинг RW. Ограничение воздушного потока, астма и специфический белок теплового шока Chlamydia pneumoniae 60.Ann Allergy, Asthma Immunol. 2008. 101 (6): 614–8.
Артикул Google Scholar
Smith-Norowitz TA, Loeffler J, Huang Y, Klein E, Norowitz YM, Hammerschlag MR, et al. Chlamydia pneumoniae Уровни антител к иммуноглобулину Е у пациентов с астмой по сравнению с пациентами, не страдающими астмой. Гелион. 2020; 6 (2): e03512.
PubMed PubMed Central Статья Google Scholar
Редеке В., Далхофф К., Бонет С., Браун Дж., Маасс М. Взаимодействие Chlamydia pneumoniae и альвеолярных макрофагов человека: инфекция и воспалительная реакция. Am J Respir Cell Mol Biol. 1998. 19 (5): 721–7.
CAS PubMed Статья PubMed Central Google Scholar
Grayston JT, Campbell LA, Kuo CC, Mordhorst CH, Saikku P, Thorn DH, et al. Новый возбудитель респираторного тракта: Chlamydia pneumoniae штамм TWAR.J Infect Dis. 1990. 161 (4): 618–25.
CAS PubMed Статья PubMed Central Google Scholar
Джонстон С.Л., Мартин Р.Дж. Chlamydophila pneumoniae и Mycoplasma pneumoniae : роль в патогенезе астмы? Am J Respir Crit Care Med. 2005. 172 (9): 1078–89.
PubMed Статья PubMed Central Google Scholar
Ян Дж., Хупер WC, Филлипс DJ, Talkington DF.Регулирование провоспалительных цитокинов в эпителиальных клетках легких человека, инфицированных Mycoplasma pneumoniae . Заражение иммунной. 2002. 70 (7): 3649–55.
CAS PubMed PubMed Central Статья Google Scholar
Chu HW, Honor JM, Rawlinson CA, Harbeck RJ, Martin RJ. Влияние респираторной Mycoplasma pneumoniae инфекции на аллерген-индуцированную гиперреактивность бронхов и воспаление легких у мышей.Заражение иммунной. 2003. 71 (3): 1520–6.
CAS PubMed PubMed Central Статья Google Scholar
Чжон Ю.С., Йео М.С., Ким Дж. Х., Ли Х. Б., О Дж. У. Инфекция Mycoplasma pneumoniae влияет на сывороточные уровни фактора роста эндотелия сосудов и интерлейкина-5 у детей с атопией. Allergy Asthma Immunol Res. 2012; 4 (2): 92–7.
CAS PubMed Статья PubMed Central Google Scholar
Gil JC, Cedillo RL, Mayagoitia BG, Paz MD. Выделение Mycoplasma pneumoniae от пациентов с астмой. Энн Аллергия. 1993. 70 (1): 23–5.
CAS PubMed PubMed Central Google Scholar
Можжевельник EF, Sonia Buist A, Cox FM, Ferrie PJ, King DR. Валидация стандартизированной версии опросника по оценке качества жизни при астме. Грудь. 1999. 115 (5): 1265–70.
CAS PubMed Статья Google Scholar
Можжевельник EF, О’Бирн П.М., Гайатт Г.Х., Ферри П.Дж., Кинг DR. Разработка и проверка анкеты для измерения контроля астмы 2. Eur Respir J. 1999; 14 (0903–1936): 902–7.
CAS PubMed Статья Google Scholar
Натан Р.А., Соркнесс CA, Косински М., Шац М., Ли Дж. Т., Маркус П. и др. Разработка контрольного теста астмы: исследование для оценки контроля астмы. J Allergy Clin Immunol. 2004. 113 (1): 59–65.
PubMed Статья Google Scholar
Gencay M, Rüdiger JJ, Tamm M, Solér M, Perruchoud AP, Roth M. Повышенная частота антител Chlamydia pneumoniae у пациентов с астмой. Am J Respir Crit Care Med. 2001. 163 (5): 1097–100.
CAS PubMed Статья Google Scholar
Кук П.Дж., Дэвис П., Танниклифф В., Эйрес Дж. Г., Ханиборн Д., Уайз Р. Chlamydia pneumoniae и астма. Грудная клетка. 1998. 53 (4): 254–9.
CAS PubMed PubMed Central Статья Google Scholar
Pasternack R, Huhtala H, Karjalainen J. Серология Chlamydophila (Chlamydia) pneumoniae и астма у взрослых: продольный анализ. J Allergy Clin Immunol. 2005. 116 (5): 1123–8.
PubMed Статья PubMed Central Google Scholar
Сасаки А., Оучи К., Маката Х, Хашимото К., Мацубара Т., Фурукава С. Влияние ингаляционных кортикостероидов на инфекцию Chlamydophila pneumoniae и Mycoplasma pneumoniae у детей с бронхиальной астмой.J Infect Chemother. 2009. 15 (2): 99–103.
CAS PubMed Статья PubMed Central Google Scholar
Tuuminen T, Varjo S, Ingman H, Weber T., Oksi J, Viljanen M. Распространенность Chlamydia pneumoniae и Mycoplasma pneumoniae , иммуноглобулин G и антител G и A в здоровой финской популяции согласно количественному ферментному анализу. иммуноанализы. Clin Diagn Lab Immunol. 2000. 7 (5): 734–8.
CAS PubMed PubMed Central Статья Google Scholar
Wark PAB, Johnston SL, Simpson JL, Hensley MJ, Gibson PG. Chlamydia pneumoniae реактивация иммуноглобулина А и воспаление дыхательных путей при острой астме. Eur Respir J. 2002; 20 (4): 834–40.
CAS PubMed Статья PubMed Central Google Scholar
Von Hertzen LC. Chlamydia pneumoniae и его роль в хронической обструктивной болезни легких. Ann Med. 1998. 30 (1): 27–37.
Артикул Google Scholar
Black PN, Scicchitano R, Jenkins CR, Blasi F, Allegra L, Wlodarczyk J и др. Серологические свидетельства инфекции Chlamydia pneumoniae связаны с тяжестью астмы. Eur Respir J. 2000; 15 (2): 254–9.
CAS PubMed Статья PubMed Central Google Scholar
Лайтинен К., Лаурила А.Л., Лейнонен М., Сайкку П. Реактивация инфекции Chlamydia pneumoniae у мышей путем лечения кортизоном.Заражение иммунной. 1996. 64 (4): 1488–90.
CAS PubMed PubMed Central Статья Google Scholar
Brière F, Bridon JM, Chevet D, Souillet G, Bienvenu F, Guret C, et al. Интерлейкин 10 побуждает В-лимфоциты пациентов с дефицитом IgA секретировать IgA. J Clin Invest. 1994. 94 (1): 97–104.
PubMed PubMed Central Статья Google Scholar
Патель К.К., Висенсио А.Г., Ду З, Цирилакис К., Сальва П.С., Уэбли В.С. Инфекционный Chlamydia pneumoniae связан с повышенным уровнем интерлейкина-8 и нейтрофилией дыхательных путей у детей с рефрактерной астмой. Pediatr Infect Dis J. 2010; 29 (12): 1093–8.
PubMed Статья PubMed Central Google Scholar
Hahn DL, Schure A, Patel K, Childs T., Drizik E, Webley W. Chlamydia pneumoniae -специфический IgE преобладает при астме и связан с тяжестью заболевания.PLoS ONE. 2012; 7 (4): e35945.
CAS PubMed PubMed Central Статья Google Scholar
Ким Р. Я., Хорват Дж. С., Пинкертон Дж. В., Старки М. Р., Эссилфи А. Т., Мэйолл Дж. Р. и др. МикроРНК-21 вызывает тяжелую, нечувствительную к стероидам экспериментальную астму, усиливая опосредованное фосфоинозитид-3-киназой подавление гистондеацетилазы 2. J Allergy Clin Immunol. 2017; 139 (2): 519–32.
CAS PubMed Статья PubMed Central Google Scholar
Schleich FN, Graff S, Guissard F, Henket M, Paulus V, Louis R. Астма у пожилых людей характеризуется повышенным содержанием нейтрофилов мокроты, изменчивостью калибра нижних дыхательных путей и захватом воздуха. Respir Res. 2021; 22 (1): 15.
CAS PubMed PubMed Central Статья Google Scholar
Ten BA, Van Dissel JT, Sterk PJ, Zwinderman AH, Rabe KF, Bel EH. Стойкое ограничение воздушного потока при неатопической астме у взрослых связано с серологическими доказательствами инфекции Chlamydia pneumoniae .J Allergy Clin Immunol. 2001. 107 (3): 449–54.
Артикул Google Scholar
Weiss SG, Newcomb RW, Beem MO. Легочная оценка детей после хламидийной пневмонии младенчества. J Pediatr. 1986. 108 (5 Pt 1): 659–64.
CAS PubMed Статья PubMed Central Google Scholar
Ансарин К., Абеди С., Готаслоу Р., Соруш М.Х., Габили К., Чапман К.Р.Инфекция Mycoplasma pneumoniae не связана с контролем астмы, тяжестью астмы и локализацией обструкции дыхательных путей. Int J Gen Med. 2011; 4: 1–4.
Google Scholar
Brusselle GG, Vanderstichele C, Jordens P, Deman R, Slabbynck H, Ringoet V, et al. Азитромицин для профилактики обострений тяжелой астмы (AZISAST): многоцентровое рандомизированное двойное слепое плацебо-контролируемое исследование. Грудная клетка. 2013. 68 (1468–3296 (Электронный)): 322–9.
PubMed Статья Google Scholar
Kraft M, Cassell GH, Pak J, Martin RJ. Mycoplasma pneumoniae и Chlamydia pneumoniae при астме: действие кларитромицина. Грудь. 2002. 121 (6): 1782–8.
CAS PubMed Статья Google Scholar
Блэк П.Н., Блази Ф., Дженкинс С.Р., Сикчитано Р., Миллс Г.Д., Рубинфельд А.Р. и др.Испытание рокситромицина на пациентах с астмой и серологические доказательства инфекции Chlamydia pneumoniae . Am J Respir Crit Care Med. 2001. 164 (4): 536–41.
CAS PubMed Статья Google Scholar
Хан Д.Л., Уэбли В. Хроническая Chlamydia pneumoniae легочная инфекция: игнорируемое объяснение эффектов макролидов при хрипе и астме? Ланцет Респир Мед. 2016; 4 (3): e8.
PubMed Статья Google Scholar
Sutherland ER, King TS, Icitovic N, Ameredes BT, Bleecker E, Boushey HA, et al. Испытание кларитромицина для лечения астмы с недостаточным контролем. J Allergy Clin Immunol. 2010. 126 (4): 747–53.
CAS PubMed PubMed Central Статья Google Scholar
Гибсон П.Г., Ян И.А., Апхэм Дж. У., Рейнольдс П. Н., Ходж С., Джеймс А. Л. и др. Влияние азитромицина на обострения астмы и качество жизни у взрослых с персистирующей неконтролируемой астмой (AMAZES): рандомизированное двойное слепое плацебо-контролируемое исследование.Ланцет. 2017; 390 (0140–6736 (Linking)): 659–68.
CAS PubMed Статья Google Scholar
Хламидийные пневмонии: обзор, патофизиология, эпидемиология
Burillo A, Bouza E. Chlamydophila pneumoniae. Инфекция Dis Clin North Am . 2010 24 марта (1): 61-71. [Медлайн].
Crosse BA. Пситтакоз: клинический обзор. J Заразить . 1990 21 ноября (3): 251-9. [Медлайн].
Джейн С., Self WH, Wunderink RG и др., Для исследовательской группы CDC EPIC. Внебольничная пневмония среди взрослых в США, требующая госпитализации. N Engl J Med . 2015 30 июля. 373 (5): 415-27. [Медлайн]. [Полный текст].
Эвиг С., Торрес А. Является ли Chlamydia pneumoniae важным патогеном у пациентов с внебольничной пневмонией ?. Eur Respir J . 2003 май. 21 (5): 741-2. [Медлайн]. [Полный текст].
Cilloniz C, Ewig S, Polverino E, et al.Микробная этиология внебольничной пневмонии и ее отношение к степени тяжести. Грудь . 2011 Апрель 66 (4): 340-6. [Медлайн]. [Полный текст].
Blasi F, Tarsia P, Aliberti S. Chlamydophila pneumoniae. Clin Microbiol Infect . Январь 2009. 15 (1): 29-35. [Медлайн].
Dumke R, Schnee C, Pletz MW и др., Для исследовательской группы Capnetz Study Group. Mycoplasma pneumoniae и Chlamydia spp. инфекция при внебольничной пневмонии, Германия, 2011-2012 гг. Emerg Infect Dis . 2015 21 марта (3): 426-34. [Медлайн]. [Полный текст].
Choroszy-Krol I, Frej-Madrzak M, Jama-Kmiecik A, Sarowska J, Gosciniak G, Pirogowicz I. Заболеваемость Chlamydophila pneumoniae у детей в 2007-2010 гг. Ад Эксп Мед Биол . 2013. 788: 83-7. [Медлайн].
Арнольд Ф.В., Саммерсгилл Дж. Т., Ладжой А.С. и др., Для исследователей Организации внебольничной пневмонии (CAPO). Мировая перспектива атипичных возбудителей внебольничной пневмонии. Am J Respir Crit Care Med . 15 мая 2007 г. 175 (10): 1086-93. [Медлайн]. [Полный текст].
Fajardo KA, Zorich SC, Voss JD, Thervil JW. Вспышка пневмонии, вызванная Chlamydophila pneumoniae, среди курсантов Академии ВВС США, Колорадо, США. Emerg Infect Dis . 2015, 21 июня (6): 1049-51. [Медлайн]. [Полный текст].
Центры по контролю и профилактике заболеваний. Сборник мер по борьбе с инфекцией Chlamydia psittaci среди людей (орнитоз) и домашних птиц (птичий хламидиоз), 2000 г. MMWR Recomm Rep . 2000 14 июля. 49: 3-17. [Медлайн]. [Полный текст].
Центры по контролю и профилактике заболеваний. Сводка болезней, подлежащих уведомлению – США, 2010 г. MMWR Morb Mortal Wkly Rep . 2012 июн 1. 59 (53): 1-111. [Медлайн]. [Полный текст].
Tipple MA, Beem MO, Saxon EM. Клиническая характеристика афебрильной пневмонии, связанной с инфекцией Chlamydia trachomatis, у детей младше 6 месяцев. Педиатрия . 1979 Февраль 63 (2): 192-7. [Медлайн].
Edelman RR, Hann LE, Simon M. Chlamydia trachomatis pneumonia у взрослых: внешний вид на рентгенограмме. Радиология . 1984 августа 152 (2): 279-82. [Медлайн].
Sharma L, Losier A, Tolbert T, Dela Cruz CS, Marion CR. Атипичная пневмония: обновленная информация о Legionella, Chlamydophila и Mycoplasma pneumonia. Clin Chest Med . 2017 Март 38 (1): 45-58. [Медлайн]. [Полный текст].
Choroszy-Krol I, Frej-Madrzak M, Hober M, Sarowska J, Jama-Kmiecik A. Инфекции, вызванные Chlamydophila pneumoniae. Адв Клин Экспер Мед . 2014 янв-фев. 23 (1): 123-6. [Медлайн]. [Полный текст].
Gacouin A, Revest M, Letheulle J, et al. Отличительные особенности внебольничной пневмонии (ВП), вызванной Chlamydophila psittaci, и ВП, вызванной Legionella pneumophila, поступившей в отделение интенсивной терапии (ОИТ). Eur J Clin Microbiol Infect Dis .2012 31 октября (10): 2713-8. [Медлайн].
Cunha BA. Атипичные пневмонии: клинический диагноз и значение. Clin Microbiol Infect . 2006 май. 12 Дополнение 3: 12-24. [Медлайн].
[Рекомендации] Baron EJ, Miller JM, Weinstein MP, et al. Руководство по использованию микробиологической лаборатории для диагностики инфекционных заболеваний: рекомендации Американского общества инфекционных болезней (IDSA) и Американского общества микробиологии (ASM) 2013 г. (a). Клиническая инфекция . 57 августа 2013 г. (4): e22-e121. [Медлайн]. [Полный текст].
She RC, Thurber A, Hymas WC, et al. Ограниченная полезность посева на Mycoplasma pneumoniae и Chlamydophila pneumoniae для диагностики инфекций дыхательных путей. Дж. Клин Микробиол . 2010 Сентябрь 48 (9): 3380-2. [Медлайн]. [Полный текст].
Бенитес А.Дж., Турман К.А., Диаз М.Х., Конклин Л., Кендиг Н.Е., Винчелл Дж. М.. Сравнение ПЦР в реальном времени и микроиммунофлуоресцентного серологического анализа для выявления инфекции chlamydophila pneumoniae в ходе расследования вспышки. Дж. Клин Микробиол . 2012 января 50 (1): 151-3. [Медлайн]. [Полный текст].
Kumar S, Hammerschlag MR. Острая респираторная инфекция, вызванная Chlamydia pneumoniae: современное состояние методов диагностики. Клиническая инфекция . 2007 15 февраля. 44 (4): 568-76. [Медлайн].
Смит К.А., Брэдли К.К., Стобирски М.Г., Тенгельсен Л.А.; Национальная ассоциация государственных ветеринаров общественного здравоохранения Комитет по сборнику пситтакозов. Сборник мер по борьбе с инфекцией Chlamydophila psittaci (ранее Chlamydia psittaci) среди людей (орнитоз) и домашних птиц, 2005 г. J Am Vet Med Assoc . 2005 15 февраля. 226 (4): 532-9. [Медлайн].
Виллегас Э., Сорлозано А., Гутьеррес Дж. Серологическая диагностика инфекции Chlamydia pneumoniae: ограничения и перспективы. J Med Microbiol . 2010 ноябрь 59 (Pt 11): 1267-74. [Медлайн].
Littman AJ, Джексон Лос-Анджелес, White E, Thornquist MD, Gaydos CA, Vaughan TL. Межлабораторная надежность теста микроиммунофлуоресценции для измерения титров антител к Chlamydia pneumoniae-специфическим иммуноглобулинам A и G. Clin Diagn Lab Immunol . 2004 г., май. 11 (3): 615-7. [Медлайн]. [Полный текст].
[Рекомендации] Mandell LA, Wunderink RG, Anzueto A, et al, от Общества инфекционных болезней Америки и Американского торакального общества. Общество инфекционных болезней Америки / Американское торакальное общество согласовали руководящие принципы ведения внебольничной пневмонии у взрослых. Клиническая инфекция . Март 2007. 44: Приложение 2: S27-72. [Медлайн]. [Полный текст].
[Рекомендации] Доуэлл С.Ф., Peeling RW, Боман Дж. И др., Для C.pneumoniae Участники семинара. Стандартизация анализов на Chlamydia pneumoniae: рекомендации Центров по контролю и профилактике заболеваний (США) и Лабораторного центра контроля заболеваний (Канада). Клиническая инфекция . 2001 15 августа. 33 (4): 492-503. [Медлайн]. [Полный текст].
Пирс В.М., Элкан М., Лит М., Макгоуэн К.Л., Ходинка Р.Л. Сравнение системы FilmArray Idaho Technology с ПЦР в реальном времени для обнаружения респираторных патогенов у детей. Дж. Клин Микробиол .2012 Февраль 50 (2): 364-71. [Медлайн]. [Полный текст].
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. FDA расширяет использование респираторной панели FilmArray. Новости и события . 15 мая 2012 г. Доступно по адресу http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm304177.htm.
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA). Выпуск новостей. FDA расширяет использование респираторной панели FilmArray. Доступно по адресу http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm304177.htm. Доступ: 26 февраля 2013 г.
Дыхательная панель BioFire Diagnostics Inc. FilmArray. Доступно по адресу http://www.biofiredx.com/pdfs/FilmArray/InfoSheet,%20FilmArray%20Respiratory%20Panel-0229.pdf. Доступ: 12 августа 2015 г.
Hvidsten D, Halvorsen DS, Berdal BP, Gutteberg TJ. Диагностика Chlamydophila pneumoniae: важность методологии в отношении времени отбора проб. Clin Microbiol Infect . Январь 2009. 15 (1): 42-9.[Медлайн].
Basarab M, Macrae MB, Curtis CM. Атипичная пневмония. Curr Opin Pulm Med . 2014 май. 20 (3): 247-51. [Медлайн].
[Рекомендации] Balsamo G, Maxted AM, Midla JW, et al. Сборник мер по борьбе с инфекцией Chlamydia psittaci среди людей (пситтакоз) и домашних птиц (птичий хламидиоз), 2017. J Avian Med Surg . 2017 Сентябрь 31 (3): 262-82. [Медлайн]. [Полный текст].
[Рекомендации] Отдел профилактики ЗППП, Национальный центр профилактики ВИЧ / СПИДа, вирусных гепатитов, ЗППП и туберкулеза, Центры по контролю и профилактике заболеваний.Руководство по лечению заболеваний, передающихся половым путем (ЗППП), 2015 г. Центры по контролю и профилактике заболеваний. 4 июня 2015 г. Доступно по адресу https://www.cdc.gov/std/tg2015/chlamydia.htm. Дата обращения: 10 ноября 2107 г..
Kohlhoff SA, Huband MD, Hammerschlag MR. Активность нового ингибитора ДНК-гиразы AZD0914 против Chlamydia trachomatis и Chlamydia pneumoniae in vitro. Противомикробные агенты Chemother . 2014 декабрь 58 (12): 7595-6. [Медлайн]. [Полный текст].
Biedenbach DJ, Huband MD, Hackel M, de Jonge BL, Sahm DF, Bradford PA.In vitro активность AZD0914, нового ингибитора бактериальной ДНК-гиразы / топоизомеразы IV, против клинически значимых грамположительных и привередливых грамотрицательных микроорганизмов. Противомикробные агенты Chemother . 2015 Октябрь 59 (10): 6053-63. [Медлайн]. [Полный текст].
van Rensburg DJ, Perng RP, Mitha IH, et al. Эффективность и безопасность немоноксацина по сравнению с левофлоксацином при внебольничной пневмонии. Противомикробные агенты Chemother . 2010 г., 54 (10): 4098-106. [Медлайн].[Полный текст].
Лю И, Чжан И, Ву Дж и др. Рандомизированное двойное слепое многоцентровое исследование фазы II, сравнивающее эффективность и безопасность перорального немоноксацина и перорального левофлоксацина при лечении внебольничной пневмонии. J Microbiol Immunol Infect . 2017 Декабрь 50 (6): 811-20. [Медлайн]. [Полный текст].
Kohlhoff SA, Hammerschlag MR. Лечение хламидийных инфекций: обновление 2014 г. Экспертное мнение Pharmacother .2015 16 февраля (2): 205-12. [Медлайн].
Роблин PM, Hammerschlag MR. Микробиологическая эффективность азитромицина и чувствительность к азитромицину изолятов Chlamydia pneumoniae от взрослых и детей с внебольничной пневмонией. Противомикробные агенты Chemother . 1998, январь, 42 (1): 194-6. [Медлайн]. [Полный текст].
Hammerschlag MR, Роблин PM. Микробиологическая эффективность левофлоксацина для лечения внебольничной пневмонии, вызванной Chlamydia pneumoniae. Противомикробные агенты Chemother . 2000 Май. 44 (5): 1409. [Медлайн]. [Полный текст].
Эларгуби А., Верховен П.О., Граттард Ф., Стефан Дж.Л., Ричард О., Поццетто Б. Острый энцефалит, связанный с респираторной инфекцией, вызванной Chlamydophila pneumoniae. Медицинская инфекция . 2013 авг. 43 (8): 345-9. [Медлайн].
Керн Дж. М., Маасс В., Маасс М. Молекулярный патогенез хронической инфекции Chlamydia pneumoniae: краткий обзор. Clin Microbiol Infect .Январь 2009. 15 (1): 36-41. [Медлайн].
Gillissen A, Paparoupa M. Воспаление и инфекции при астме. Клин Респир Дж . 2015 июл.9 (3): 257-69. [Медлайн].
Ивен М.М., Хойманс В.Ю. Вовлечение Chlamydia pneumoniae в атеросклероз: дополнительные доказательства отсутствия доказательств. Дж. Клин Микробиол . 2005, январь, 43 (1): 19-24. [Медлайн].
Andraws R, Berger JS, Brown DL. Влияние антибактериальной терапии на исходы пациентов с ишемической болезнью сердца: метаанализ рандомизированных контролируемых исследований. JAMA . 1 июня 2005 г. 293 (21): 2641-7. [Медлайн]. [Полный текст].
Grayston JT. Инфекции, вызванные штаммом Chlamydia pneumoniae TWAR. Клиническая инфекция . 1992 15 ноября (5): 757-61. [Медлайн].
Fine MJ, Smith MA, Carson CA, et al. Прогноз и исходы пациентов с внебольничной пневмонией. Метаанализ. JAMA . 1996, 10 января. 275 (2): 134-41. [Медлайн].
Файл TM Jr, Tan JS, Plouffe JF.Роль атипичных возбудителей: Mycoplasma pneumoniae, Chlamydia pneumoniae и Legionella pneumophila в респираторной инфекции. Инфекция Dis Clin North Am . 1998 Сентябрь 12 (3): 569-92, vii. [Медлайн].
Marrie TJ, Peeling RW, Reid T., De Carolis E. Виды хламидий как причина внебольничной пневмонии в Канаде. Eur Respir J . 2003 май. 21 (5): 779-84. [Медлайн]. [Полный текст].
Cipriano A, Machado A, Santos FV, Abreu MA, Castro RS.[Пситтакоз человека: отчет о болезни]. Порт Acta Med . 2019 28 февраля. 32 (2): 161-4. [Медлайн].
Kohlhoff SA, Huerta N, Hammerschlag MR. In vitro активность омадациклина против Chlamydia pneumoniae . Противомикробные агенты Chemother . 2019 Февраль 63 (2): [Medline]. [Полный текст].
Chlamydia pneumoniae Белок GroEL1 связан с клеточной поверхностью и необходим для инфицирования клеток HEp-2
РЕЗЮМЕ
Chlamydia pneumoniae – важный облигатный внутриклеточный патоген, который реплицируется внутри включения в эукариотической клетке.Первоначальным событием хламидийной инфекции является присоединение и последующее поглощение инфекционных элементарных тел (ЭТ) клеткой человека. Для этих процессов требуются еще не идентифицированные поверхностные белки бактерий и эукариот. Белок GroEL1, который проявляет очень сильную антигенность и in vitro может активировать различные эукариотические клетки, является потенциальным фактором патогенности. Мы локализовали белок в процессе заражения и обнаружили его во включении, но вне хламидийных частиц.GroEL1 также был локализован на поверхности EB, и белок мог смываться с EB. Латексные шарики, покрытые рекомбинантно продуцированным GroEL1 (rGroEL1), связывались дозозависимым образом с клетками HEp-2. Аналогичным образом, GroEL1, когда экспрессируется и отображается на поверхности дрожжевых клеток, опосредует адгезию к клеткам HEp-2. Интересно, что гомологичные белки GroEL2 и GroEL3 не проявляли адгезионных свойств. Инкубация первичных эндотелиальных клеток пупочной вены с растворимыми латексными шариками, покрытыми GroEL1 и GroEL1, активировала транслокацию общего фактора транскрипции NF-κB в ядро.Наконец, предварительная инкубация клеток HEp-2 с rGroEL1 значительно снизила последующее инфицирование C. pneumoniae , хотя адгезия инфекционных бактерий к эукариотическим клеткам не пострадала. Взятые вместе, эти данные подтверждают роль внеклеточного GroEL1 в возникновении хламидийной инфекции.
Человеческий патоген Chlamydia pneumoniae – облигатная внутриклеточная грамотрицательная бактерия, характеризующаяся уникальным двухфазным жизненным циклом. Инфекционные, но метаболически неактивные элементарные тельца (EB) прикрепляются к эукариотическим клеткам и вторгаются в них, уклоняются от слияния с лизосомами и дифференцируются в метаболически активные сетчатые тельца внутри вакуоли хозяина (называемой включением), в которой они размножаются.После периода роста сетчатые тела повторно дифференцируются в EB, которые высвобождаются из инфицированных клеток (25, 30). Хламидии проникают в клетки-хозяева разными путями, и считается, что эта универсальность объясняет видовые различия и тропизм тканей (63).
C. pneumoniae вызывает несколько тяжелых заболеваний дыхательных путей (42, 46, 79). Кроме того, сообщалось о связи между инфекцией C. pneumoniae , атеросклерозом и ишемической болезнью сердца (64, 86).В последнее время патоген также был связан с несколькими другими хроническими заболеваниями (23, 26, 52, 77, 89).
Молекулярные механизмы патогенеза хламидий плохо изучены, и многие факторы, которые вносят вклад в патогенность этой бактерии, еще не определены точно. Заражение EB требует как прикрепления к клеткам-мишеням, так и поглощения ими. Несколько отчетов продемонстрировали, что гликозаминогликаны (ГАГ) на клетках-хозяевах участвуют в прикреплении и последующей инфекции, но остаются разногласия относительно точных задействованных механизмов (90; рассмотрено в ссылке 10).Однако недавно было показано, что белок OmcB, связанный с бактериальной поверхностью, является основным GAG-связывающим адгезином (19, 55, 76). Другие бактериальные белки, включая богатые цистеином мембранные белки Hsp70 и MOMP, были проанализированы на предмет их потенциального вклада в прикрепление EB Chlamydia trachomatis и Chlamydia psittaci к эукариотическим клеткам (47, 60, 70, 76, 78, 80), но ни один из них не действует как хламидийный поверхностный лиганд с высоким сродством. Интересно, что антитела, направленные против PmpD из C.trachomatis или Pmp21 из C. pneumoniae значительно снижали последующую инфекцию, предполагая, что они могут представлять хламидийные адгезины (13, 85). Помимо GAG, компоненты хозяина, которые участвуют в связывании EB, до сих пор ускользали от обнаружения, хотя в этом процессе участвовала протеин-дисульфид-изомераза (11, 14). Таким образом, молекулярные механизмы, участвующие в прикреплении и проникновении хламидий, остаются в значительной степени нерешенными и противоречивыми.
Одним из немногих белков, которые до сих пор идентифицированы как имеющие отношение к хламидийному патогенезу, является белок GroEL1, также называемый белком теплового шока 60 (Hsp60) (84).Этот белок относится к шаперонинам группы I, продуцируемым почти всеми прокариотическими и эукариотическими клетками, которые, как внутриклеточные белки, помогают в правильном сворачивании растущих или денатурированных белков как в нормальных, так и в стрессовых условиях (8, 91). Однако очевидно, что молекулярные шапероны имеют дополнительные функции. В нескольких сообщениях указано, что молекулярные шапероны, продуцируемые патогенными бактериями, такими как Hsp60, могут действовать как внутриклеточные, поверхностные или внеклеточные сигналы в ходе инфекционных процессов (обзор см. В ссылке 27).И наоборот, клетки человека реагируют на инфекцию или другие формы стресса, секретируя белки теплового шока (20, 58, 73).
Анализ последовательности генома показал, что все виды хламидий обладают тремя groEL -подобными генами ( groEL1 , groEL2 и groEL3 ) (36, 62, 75). GroEL1 демонстрирует наивысшую гомологию с белками GroEL из других организмов, а функция шаперонина хламидийного GroEL1 была продемонстрирована путем комплементации чувствительного к температуре мутанта GroEL Escherichia coli (3, 37).
Рекомбинантный C. pneumoniae GroEL1 (rGroEL1) может стимулировать клетки врожденного иммунитета in vitro через Toll-подобный рецептор 2 (TLR2) и TLR4 и, таким образом, может способствовать провоспалительным ответам, наблюдаемым во время инфекции (12). Более того, C. pneumoniae GroEL1 может активировать гранулоцитарно-макрофагальный колониестимулирующий фактор в эпителиальных клетках бронхов человека почти так же эффективно, как жизнеспособные или УФ-инактивированные целые бактерии, вызывая устойчивый провоспалительный фенотип (43).Точно так же C. trachomatis GroEL1 индуцирует эндотелиальные клетки, гладкомышечные клетки и макрофаги для выработки факторов адгезии и провоспалительных цитокинов. В обоих случаях GroEL1 действует путем активации NF-κB (9, 40, 51, 66, 81).
Иммунные ответы на хламидийный GroEL1 значительно коррелируют с последствиями заболевания у людей, и от 80 до 90% пациентов, инфицированных C. trachomatis , имеют антитела, направленные против GroEL1 (7, 17, 28, 65). Но до сих пор неясно, вносят ли специфические антитела к хламидийному GroEL1 прямой вклад в патогенез заболевания.Предполагается, что в ходе хламидийных инфекций бактериальный GroEL вызывает перекрестно-реактивные иммунные ответы на эндогенный GroEL, и это может составлять большую часть повреждений тканей, наблюдаемых при хронических инфекциях (57, 91). Однако на морских свинках было показано, что иммунизация нативным GroEL1 фактически снижает степень крупной глазной патологии, вызванной последующей инфекцией Chlamydophila caviae (GPIC) , что указывает на то, что ответ против GroEL1 обеспечивает определенную степень защиты от глазных болезней. болезнь (59).
Высокая степень антигенности GroEL1 у пациентов означает, что белок легко доступен для иммунной системы, возможно, потому, что он локализован на поверхности хламидийных частиц. Ранние исследования изолированных комплексов внешней мембраны из C. trachomatis и C. psittaci EBs действительно указали на возможность того, что GroEL1 может быть ассоциирован с хламидийными мембранами (3). В настоящем исследовании мы показываем, что C. pneumoniae GroEL1 присутствует на поверхности инфекционных EB и связывается с клетками-хозяевами.Кроме того, мы демонстрируем, что предварительная инкубация клеток-мишеней с хламидийным rGroEL1 существенно подавляет процесс инфицирования.
МАТЕРИАЛЫ И МЕТОДЫ
Эукариотические клетки. Клетки Hep-2 (ATCC CCL-23) размножали, как описано ранее (33). Для иммунофлуоресцентных анализов клетки высевали на покровные стекла (диаметром 1 см) и инкубировали в течение 48 часов при 37 ° C. Клетки, которые будут использоваться для хламидийной инфекции, высевали в 25-см полистирольные колбы 2 с образованием сливающихся монослоев.Анализы прикрепления проводили с использованием аликвот (1 × 10 5 ) клеток HEp-2, высеянных на покровные стекла за 24 ч до экспериментов.
Первичные эндотелиальные клетки пупочной вены человека (HUVEC) (предоставлены J. Schrader, Institut fuer Kreislaufphysiologie, HHU Duesseldorf) культивировали при 37 ° C (второй-третий пассаж) в среде Хэма F12K (Invitrogen) с добавлением 10% эмбриона крупного рогатого скота. сыворотка, 30 мкг / мл добавки для роста эндотелиальных клеток (Sigma), 10 Ед / мл гепарина (Sigma) и 6% CO 2 .
Штаммы бактерий и дрожжей. Chlamydia pneumoniae Респираторный изолят GiD культивировали, как описано ранее (32). Escherichia coli XL-1 Blue (Stratagene) выращивали в LB с 50 мкг / мл ампициллина или без него.
Saccharomyces cerevisiae CEN.PK2-1C ( MAT leu2-3 , 112 ura3-52 trp1-289 his3-Δ1 MAL2-8 c SUC2 ) (18) выращивали в среде YP с добавлением 4 мг / л аденина и 20 мг / л триптофана плюс 2% глюкозы (YPD) или в синтетической среде плюс 2% глюкозы (SD) (72 ).Штамм EBY100 S. cerevisiae ( MAT ura 3- 52 trp1 leu2 Δ 1 his3 Δ 200 pep 4 :: HIS3 prb1 Δ 1.6R can1 GAL ), используемый для экспериментов по адгезии, выращивали в синтетической среде, содержащей 2% рафинозы. (SR) или 2% галактозы (SG) (72) (Invitrogen) (55). Для экспрессии слитых белков Aga2p или Aga2p в штамме EBY100 клетки выращивали в соответствии с руководством, предоставленным Invitrogen.
Плазмиды.Плазмида pMal-c2 (New England Biolabs) была модифицирована для обеспечения возможности амплификации в дрожжах путем добавления фрагмента ДНК S. cerevisiae CEN6 / ARSh5 / URA3 длиной 1921 п.н., , который был амплифицирован из плазмиды pRS316 (74) с помощью ПЦР с использованием олигонуклеотидов. C-284 и C-285 (таблица 1) и полимераза Pfx (Invitrogen). Продукт ПЦР интегрировали в расщепленную SwaI плазмиду pMal-c2 путем гомологичной рекомбинации в дрожжах с использованием протокола трансформации, описанного Schiestl и Gietz (69). Затем была выделена общая дрожжевая ДНК (35) и использована для трансформации E.coli для восстановления рекомбинантной плазмиды pAC-2. Слитый белок мальтозо-связывающий белок (MBP) -инвазин (Inv) экспрессировали из плазмиды pRI284 (любезно предоставленной П. Дершем) (16).
Клонирование и экспрессия генов groEL Гены C. pneumoniae groEL2 (Cpn0777) и groEL3 (Cpn0898) амплифицированы с помощью ПЦР с использованием пар праймеров C-123 / C-124 и C-125 / C. -126 соответственно (таблица 1) и C. pneumoniae геномная ДНК в качестве матрицы.Оба ампликона были клонированы в расщепленную EcoRI дрожжевую E. coli челночную плазмиду pAC-2 путем гомологичной рекомбинации в S. cerevisiae . Интеграция гена groEL2 или groEL3 в pAC-2 привела к слиянию с геном malE для MBP. Ген groEL из E. coli K-12 и фрагмент гена, кодирующий адгезионный домен Inv из Yersinia pseudotuberculosis (п.н. с 2235 по 2826), были амплифицированы с использованием олигонуклеотидов C-154 / C-155 и C-578 / С-579 соответственно (таблица 1).Ампликоны трансформировали в дрожжи вместе с плазмидой pAC-2, расщепленной BglII / EcoRI, с получением плазмиды экспрессии GroEL или Inv, кодирующей слитые белки, меченные на N-конце His 6 .
Экспрессия белка была индуцирована в E. coli , выращенных в жидкой культуре, обработкой 1 мМ IPTG (изопропил-β-d-тиогалактопиранозид) в течение 5 часов. Экспрессию и очистку слитых белков MalE MBP-Inv, MBP-LacZ, MBP-GroEL2 и MBP-GroEL3 проводили, как было предложено поставщиком плазмиды-предшественника pMal-c2 (New England Biolabs).Меченный His rGroEL1 из C. pneumoniae , GroEL из E. coli и белок Inv из Y. pseudotuberculosis были экспрессированы и очищены, как описано ранее (33). GroEL и GroEL1 выделяли в присутствии 20 мкг / мл полимиксина B (Sigma) (33). Очищенные белки трижды диализовали против фосфатно-солевого буфера (PBS) и хранили при -20 ° C. Остаточный липополисахарид (ЛПС) удаляли пропусканием через колонки с полимиксином B-агарозой (предварительно упакованные колонки Detoxi-Gel AffinityPak; каталожный номер.20344, Pierce) согласно рекомендациям производителя. Концентрация эндотоксина в этих белковых препаратах составляла <0,05 единиц эндотоксина / мг белка, как определено анализом лизата амебоцитов Limulus (анализы проводили в соответствии со стандартами FDA ACILA AG, Mörfelden-Walldorf, Германия).
Для экспрессии GroEL1 на поверхности дрожжевых клеток полноразмерную кодирующую последовательность groEL1 амплифицировали с помощью ПЦР с использованием олигонуклеотидов C-236 и C-237 (таблица 1) с C.pneumoniae GiD и клонировали в расщепленный EcoRI / NotI вектор pYD1 (Invitrogen) путем гомологичной рекомбинации в S. cerevisiae . Плазмиды, экспрессирующие Aga2-Inv и Aga2-OmcB, были описаны Mölleken и Hegemann (55). Все использованные конструкции были проверены секвенированием.
Иммунолокализация белков C. pneumoniae . Клетки Hep-2, выращенные на стеклянных покровах в 24-луночных планшетах в течение 48 ч (1 × 10 5 на лунку), инфицировали C.pneumoniae GiD при множественности инфекции (MOI) 0,5. Через 60 часов после инфицирования клетки промывали один раз PBS, фиксировали 3,7% формальдегидом в течение 1 часа и повышали проницаемость PBS, содержащим 0,5% Triton X-100, или фиксировали 96% метанолом в течение 10 минут. Затем монослой трижды промывали PBS (39). Поликлональные антитела, направленные против OmpA (1:20) или белка C. trachomatis, S1 (1: 4), или моноклональные антитела против DnaK (1:50) были любезно предоставлены S. Birkelund и F.Вупперманн (F. Wuppermann и J. Hegemann, неопубликовано) (39, 50). Поликлональные антитела, направленные против C. pneumoniae OmcB, были предварительно абсорбированы против белка rOmcB (1:25) (55). Моноклональное антитело против GroEL1 (A57B9) (1: 200) направлено против N-концевого эпитопа GroEL1 и было любезно предоставлено R. Morrison (88). Вторичные антитела, меченные флуоресцеинизотиоцианатом, использовали в разведении 1:20 (Dako). Вторичные антитела, меченные Cy3, использовали в разведении 1: 200 (Sigma).Клетки просматривали с помощью Zeiss Axioskop.
Для иммунолокализации GroEL1 на хламидийных EB, хламидии очищали с использованием градиента Перколла (2). Жизнеспособные или фиксированные метанолом EB обрабатывали для иммунофлуоресценции, как описано выше.
Фракционирование хламидийного белка GroEL1 из инфекционных EB. Хламидийные частицы (EB) очищали на градиентах Перколла центрифугированием при 30000 × g , как описано ранее (2). Бактериальный осадок (приблизительно 1 × 10 8 частиц) ресуспендировали в 250 мкл PBS, инкубировали в течение 2 часов при 4 ° C, снова осаждали при 30000 × г в течение 30 минут, инкубировали еще 2 часа при 4 ° C. ° C в 250 мкл PBS и восстанавливали центрифугированием при 30,000 × g .Осадок хламидийного EB затем готовили для электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE). Два супернатанта лиофилизировали отдельно и растворяли в буфере SDS-PAGE. Осадок и две фракции супернатанта затем подвергали SDS-PAGE и Вестерн-блоттингу с использованием антител OmpA, DnaK и GroEL1. Загруженные промывные фракции содержали в три раза больше белка, чем образцы осадка.
SDS-PAGE и Вестерн-блот-анализ. Белки фракционировали SDS-PAGE и подвергали Вестерн-блот-анализу.Антитело, специфичное для метки His (Qiagen), использовали для обнаружения rGroEL1 с меткой His. В качестве вторичных антител использовали антитела против мыши или кролика, каждое из которых связано с щелочной фосфатазой (Promega).
Анализы адгезии с покрытыми белком латексными шариками. Латексные шарики (Sigma) (диаметр, 1 мкм) были покрыты белком (16). Эффективность покрытия была проанализирована с помощью иммунофлуоресцентной микроскопии и иммуноблоттинга и составила от 90 до 95%. Анализы адгезии с покрытыми белком латексными шариками и клетками HEp-2 проводили, как сообщалось ранее (16), и определяли среднее количество шариков, связанных с клетками.
Анализы адгезии дрожжей. Анализы адгезии дрожжей проводили с клетками HEp-2, культивированными на стеклянных покровных стеклах, как описано ранее (55). Вкратце, дрожжевые клетки (1 × 10 6 в 1 мл PBS), экспрессирующие только Aga2p или специфический слитый белок на клеточной поверхности, добавляли к 1 × 10 5 клеток HEp-2. Покровные стекла инкубировали 1 ч. при 4 ° C при осторожном встряхивании, трижды промывали 0,5 мл PBS и фиксировали 3,7% формальдегидом. Под микроскопом подсчитывали количество дрожжевых клеток, прикрепленных к 1000 клеткам HEp-2.Каждый эксперимент повторяли четыре раза.
Анализы активации NF-κB. HUVEC высевали на покрытые желатином 24-луночные планшеты с плотностью 2 × 10 5 клеток / лунку и оставляли на 24 часа при 37 ° C в присутствии 6% CO. 2 . Клетки инкубировали либо с белком, либо с шариками, покрытыми белком. В качестве контроля HUVEC были инфицированы C. pneumoniae (MOI, ∼10) и проанализированы через 2 часа. Субклеточную локализацию NF-κB анализировали в фиксированных клетках с использованием поликлонального кроличьего антитела p65 против человеческого NF-κB (Santa Cruz Biotechnology) и вторичного антитела, меченного флуоресцеинизотиоцианатом (Sigma), и процент HUVEC, демонстрирующих сильную ядерную активность. сигнал был определен.
Анализ конкуренции на инфекцию. Конфлюентные монослои клеток HEp-2, выращенные на покровных стеклах, промывали PBS и инкубировали с 20 или 200 мкг / мл rGroEL1 или бычьим сывороточным альбумином (BSA) в течение 2 часов при 37 ° C при осторожном перемешивании. Очищенные в градиенте перколла C. pneumoniae добавляли EB и инкубировали еще 2 часа при 37 ° C при легком перемешивании. Клетки HEp-2 промывали три раза, покрывали хламидийной средой для выращивания и инкубировали в течение 3 дней при 37 ° C. Клетки фиксировали и хламидийные включения метили с использованием хламидий-специфических антител, направленных против ЛПС (Pathfinder; Bio-Rad).Количество включений в 80 полях микроскопа (20 / покровное стекло) определяли и выражали как процент от количества, обнаруженного в образцах, обработанных PBS.
Проточный цитометрический анализ адгезии CFSE-меченных C. pneumoniae EB к человеческим клеткам Очищенные C. pneumoniae EB (2 × 10 8 ) метили в течение 1 часа при 37 ° C 25 мкмоль карбоксифлуоресцеин сукцинимидил сложного эфира (CFSE) (Molecular Probes) и дважды промывали PBS, содержащим 1% BSA, как описано ранее (71).Конфлюэнтные монослои клеток HEp-2 или HUVEC в 24-луночных планшетах инкубировали в течение 2 ч при 37 ° C с 20 мкг / мл или 200 мкг / мл рекомбинантного белка или 500 мкг / мл гепарина и CFSE-меченного C. pneumoniae. Затем добавляли EB (MOI, 10) и оставляли на 1 час. Клетки промывали PBS, трипсинизировали и фиксировали формальдегидом, а степень бактериальной адгезии измеряли проточной цитометрией с использованием FACSAria (BD Biosciences).
РЕЗУЛЬТАТЫ
C. pneumoniae Белок GroEL1 локализован внутри хламидийного включения, но вне бактериальной клетки.Чтобы определить локализацию белка GroEL1 во время хламидийной инфекции, эпителиальные клетки человека HEp-2 инфицировали C. pneumoniae . Через 60 часов после инфицирования инфицированные клетки фиксировали либо метанолом (делая все хламидийные антигены доступными для антител), либо формальдегидом и тритоном X-100 (внутрихламидийные антигены остаются недоступными) и обрабатывали для непрямой иммунофлуоресценции (39). В фиксированных метанолом клетках как белки внешней мембраны хламидии (OmcB и OmpA), так и внутриклеточные хламидийные белки DnaK и S1 могут быть легко обнаружены в хламидийном включении (рис.1A – D; см. рис. S1B и S1J в дополнительном материале). Окрашивание включений моноклональным антителом, направленным против белка GroEL1, аналогично показало, что антиген локализован во включении хламидий (фиг. 1A и B; см. Фиг. S1F в дополнительном материале). В клетках, фиксированных формальдегидом, внехламидийные белки OmcB и OmpA все еще могут быть обнаружены внутри включения (рис. 1C и D; см. Рис. S1L в дополнительном материале), в то время как внутриклеточные хламидийные белки DnaK и S1 не были доступны для антитела в условиях Эти условия фиксации (рис.1А и В; см. рис. S1D в дополнительном материале). Обратите внимание, что формальдегид не повреждает эпитопы S1 и DnaK, о чем свидетельствует тот факт, что последующая фиксация метанолом позволяет антителам распознавать оба белка (данные не показаны). Интересно, что окрашивание хламидийных включений моноклональным антителом, направленным против GroEL1, показало, что GroEL1 может быть обнаружен внутри включения, но вне бактериальных клеток (фиг. 1A и B; см. Фиг. S1H в дополнительном материале).Тот факт, что внутрихламидийные белки DnaK и S1 не обнаруживаются в фиксированных формальдегидом клетках, указывает на то, что бактериальный лизис внутри включения незначителен. Такой же паттерн локализации наблюдали с поликлональным антителом, направленным против GroEL1 (данные не показаны). В совокупности эти данные указывают на то, что часть хламидийного белка GroEL1 секретируется хламидийными частицами, присутствующими во включении.
РИС. 1.Локализация хламидийного GroEL1 внутри хламидийного включения.Клетки HEp-2, инфицированные C. pneumoniae при MOI 0,5 в течение 60 часов, фиксировали либо метанолом для визуализации как внутри-, так и внехламидийных белков, либо формальдегидом плюс 0,5% Triton X-100 для визуализации внехламидийных белков (5). Непрямая иммунофлуоресцентная микроскопия выполнялась с использованием антител против внутрихламидийной DnaK (A и C) или C. trachomatis, рибосомного белка S1 (B и D) и локализованного на поверхности хламидийного OmcB (C и D) и хламидийного GroEL1 (A и B).Пруток, 10 мкм.
C. pneumoniae Белок GroEL1 связан с поверхностью инфекционных EB. Описанный выше паттерн локализации GroEL1 выявлялся на протяжении всего процесса инфицирования (от 24 до 72 часов после заражения) (данные не показаны). Поскольку большинство сетчатых телец внутри включений повторно дифференцировались в EB через 72 часа после инфицирования, этот результат предполагал, что инфекционные EB, высвобождаемые из инфицированных эукариотических клеток, также могут нести внеклеточный белок GroEL1.Чтобы напрямую решить эту проблему, инфекционные EB очищали на градиентах Percoll и либо окрашивали без фиксации, либо сначала фиксировали метанолом, а затем окрашивали GroEL1-специфическим моноклональным антителом. В обоих случаях на EB был обнаружен сильный сигнал флуоресценции, что указывает на то, что белок GroEL1 действительно присутствует на поверхности инфекционных EB (рис. 2A). Напротив, окрашивание нефиксированных EB с использованием антитела, направленного против хламидийного белка DnaK, не давало сигналов, в то время как белок DnaK легко обнаруживался после фиксации метанолом (рис.2А).
РИС. 2.Поверхностная локализация C. pneumoniae GroEL1. (A) Хламидийный белок GroEL1 может быть обнаружен на очищенных в градиенте Percoll EB без фиксации (без фиксации), а также после фиксации метанолом с использованием непрямой иммунофлуоресцентной микроскопии. Антитела, направленные против внутрихламидийного белка DnaK, смогли обнаружить антиген только после фиксации метанолом. Пруток, 1 мкм. (B) EB, очищенные в градиенте Percoll, дважды промывали PBS; входные, промывные фракции 1 и 2 и осадок разделяли с помощью SDS-PAGE и анализировали вестерн-блоттингом с использованием антител против хламидийных OmpA, DnaK и GroEL1.Загруженные промывные фракции содержали в три раза больше белка, чем образцы осадка. Данные представляют несколько отдельных экспериментов.
Затем мы спросили, какая часть белка GroEL1 C. pneumoniae связана с поверхностью EB. В экспериментах по фракционированию EB, очищенные в градиенте Percoll, промывали PBS и определяли количество GroEL1, присутствующего в осадке и супернатанте, с помощью вестерн-блоттинга (фиг. 2B). Менее 10% входящего белка DnaK и менее 5% контрольного белка OmpA было обнаружено в промывных растворах, в то время как примерно 50% общего белка GroEL1, обнаруженного на входе, было обнаружено в промывных растворах (рис.2B; сравните дорожку 1 с дорожками 2 и 3). Приблизительно 50% белка GroEL1 оставалось связанным с гранулированными EB (фиг. 2B, дорожка 4). Этот результат предполагает, что по крайней мере половина хламидийного белка GroEL1 связана с бактериальной поверхностью.
Латексные шарики, покрытые GroEL1, связывают клетки HEp-2. Локализация C. pneumoniae GroEL1 на поверхности инфекционных EB предполагает, что белок может взаимодействовать с эукариотическими клетками во время процесса инфицирования. Чтобы напрямую проверить эту возможность, клетки HEp-2 подвергали воздействию латексных шариков, покрытых очищенным His-меченным хламидийным белком rGroEL1 (рис.3А и В).
РИС. 3.Адгезия латексных шариков, покрытых rGroEL1, к клеткам HEp-2. (A) Окрашенный кумасси синим SDS-PAGE аффинно очищенного белка rGroEL1 после экспрессии в E. coli (полоса C) и анализа очищенного rGroEL1 методом вестерн-блоттинга с использованием антитела против N-концевой метки His (полоса WB; стрелкой отмечена полоса белка rGroEL1). Дорожка M, маркеры молекулярной массы. (B) Латексные шарики покрывали BSA или His-меченными рекомбинантными белками, как указано (концентрация белка во время покрытия 200 мкг / мл).Покрытые шарики инкубировали с клетками HEp-2. Адгезию покрытых белком латексных шариков визуализировали с помощью фазово-контрастной микроскопии (увеличение × 63). Стрелки указывают латексные шарики, прикрепленные к клеткам HEp-2. Пруток, 10 мкм. (C) Дозозависимая адгезия шариков, покрытых rGroEL1, к клеткам HEp-2. Гранулы были покрыты белком rGroEL1 с разными концентрациями. Контрольные шарики (BSA, Inv и E. coli GroEL) всегда были покрыты белком с концентрацией 200 мкг / мл. Результаты экспериментов по адгезии представлены в виде связанных шариков на клетку HEp-2 ( n = 1000 клеток HEp-2; количество экспериментов = 4).Представленные данные представляют собой средние значения ± стандартное отклонение.
Световая микроскопия показала, что контрольные латексные шарики, покрытые BSA, редко связываются с клетками HEp-2 (рис. 3B и C). В качестве положительного контроля гранулы покрывали известным бактериальным адгезином, инвазином (InV) из Yersinia pseudotuberculosis (16). Гранулы, покрытые His-tagged rInv, в большом количестве связываются с эукариотическими клетками (фиг. 3C). Интересно, что гранулы, покрытые rGroEL1 с меткой His, также показали значительную ассоциацию с клетками HEp-2 (рис.3С). В отличие от него меченный His белок rGroEL E. coli не опосредовал адгезию гранул к клеткам HEp-2 (фиг. 3C). Связывание шариков, покрытых rGroEL1 C. pneumoniae , зависело от дозы; когда шарики были покрыты rGroEL1 в концентрации 200 мкг / мл, наблюдали в среднем от 1 до 2 шариков на клетку (фиг. 3C). Эти эксперименты показывают, что белок rGroEL1 из C. pneumoniae способен прикрепляться к эукариотическим клеткам.
GroEL1 опосредует прикрепление дрожжевых клеток к клеткам HEp-2.Чтобы подтвердить результаты, полученные с помощью теста на латексных шариках, мы использовали систему дрожжевого дисплея, которая позволяет изучать взаимодействие бактериальных белков с клетками человека (55). С этой целью полноразмерный белок GroEL1 был экспрессирован в виде слияния с дрожжевой субъединицей рецептора -агглютинина Aga2p, которая заякорена в клеточной стенке дрожжей через субъединицу Aga1p. Плазмидный ген слитого белка Aga2-GroEL1 экспрессируется через индуцируемый галактозой промотор GAL1 .Aga2-GroEL1 может быть обнаружен на поверхности дрожжевых клеток с помощью антител против GroEL1 (данные не показаны). Клетки дрожжей, экспрессирующие Aga2-GroEL1, инкубировали с клетками HEp-2, засеянными на покровные стекла. В отличие от дрожжевых клеток, экспрессирующих только Aga2p, дрожжевые клетки, представляющие слитый белок Aga2-GroEL1 на клеточной поверхности, показали значительное сродство к клеткам HEp-2, подобное сродству дрожжевых клеток, экспрессирующих один из двух белков положительного контроля, белок Inv и белок C. pneumoniae OmcB (рис.4). Таким образом, эксперименты с дрожжевым дисплеем подтверждают, что C. pneumoniae GroEL1 может придавать способность прикрепляться к эпителиальным клеткам человека.
РИС. 4.Дрожжевые клетки, экспрессирующие Aga2-GroEL1 на поверхности дрожжевых клеток, прикрепляются к клеткам человека. (A) Нетрансформированные дрожжевые клетки или дрожжевые клетки, экспрессирующие Aga2, Aga2-Inv, Aga2-OmcB или Aga2-GroEL1 из плазмиды, инкубировали с 1 × 10 5 клеток HEp-2, и количество дрожжевых клеток, связанных с HEp -2 клетки определяли микроскопически.Показаны типичные микрофотографии. Пруток, 10 мкм. (B) Схематическое изображение количества дрожжевых клеток, экспрессирующих Aga2, Aga2-Inv, Aga2-OmcB и Aga2-GroEL-1, прикрепленных к 1000 клеток HEp-2 (количество экспериментов = 4). Представленные данные представляют собой средние значения ± стандартное отклонение.
GroEL1, но не GroEL2 или GroEL3, может опосредовать адгезию к клеткам человека. Мы проверили, была ли способность C. pneumoniae GroEL1 прикрепляться к клеткам HEp-2 общим свойством GroEL и других GroEL-подобных белков.Все секвенированные к настоящему времени геномы хламидий содержат три гена, подобных groEL ( groEL1 , groEL2 и groEL3 ), которые кодируют белки, которые на 20-60% идентичны прототипу белка GroEL из E. coli (36, 62, 75). GroEL1 из штамма GiD C. pneumoniae демонстрирует 33% и 24% идентичности с GroEL2 и GroEL3, соответственно (см. Рис. S2 в дополнительном материале). Чтобы проверить, все ли эти GroEL-подобные белки обладают способностью прикрепляться к клеткам HEp-2, мы экспрессировали и очищали MBP-меченый C.pneumoniae rGroEL2 и rGroEL3 и выполнили анализ гранул. Ни один из этих белков не проявлял связывания с клетками HEp-2 на уровнях, значительно превышающих уровни, обнаруженные для отрицательных контролей MBP и BSA (фиг. 5). Напротив, положительный контроль, меченый МВР белок Y. pseudotuberculosis Inv, опосредовал значительную адгезию гранул к клеткам человека (фиг. 5). Вместе эти эксперименты показывают, что из протестированных GroEL / GroEL-подобных белков GroEL1 является единственным, способным опосредовать связывание с эукариотическими клетками.
РИС. 5. Анализна связывание C. pneumoniae латексных шариков, покрытых rGroEL2 или rGroEL3, с клетками HEp-2. Латексные шарики покрывали указанными белками (концентрация белка во время покрытия 200 мкг / мл) и инкубировали с клетками HEp-2, как описано в разделе «Материалы и методы». Результаты экспериментов по связыванию представлены в виде связанных шариков на одну клетку HEp-2 ( n = 1000 клеток HEp-2; количество экспериментов = 4). Представленные данные представляют собой средние значения ± стандартное отклонение.
Белок rGroEL1 из C.pneumoniae активирует NF-κB в HUVEC. Инфекция культивируемых эндотелиальных клеток C. pneumoniae активирует повсеместный ядерный фактор NF-κB, который при перемещении из цитозоля в ядро усиливает экспрессию различных генов. участвуют в воспалительных и пролиферативных ответах (1, 15, 44). Активация NF-κB, индуцированная хламидиями, приводит к активации генов тканевого фактора, E-селектина и молекул адгезии ICAM-1 и VCAM-1 (22, 40, 44, 56).Мы спросили, является ли локализованный на поверхности GroEL1 из C. pneumoniae составляющей, активирующей NF-κB в эндотелиальных клетках. Чтобы проверить эту гипотезу, первичные HUVEC культивировали и инкубировали с C. pneumoniae , только с буфером или с rGroEL1, и определяли количество клеток, показывающих транслокацию NF-κB из цитозоля в ядро. Во всех экспериментах с использованием рекомбинантного белка концентрация эндотоксина LPS (связывание которого также приводит к ядерному захвату NF-κB) в конечных препаратах GroEL1 была <0.05 единиц эндотоксина / мг белка, как определено с помощью анализа лизата амебоцитов Limulus . Инкубация HUVEC с инфекционными EB C. pneumoniae индуцировала транслокацию NF-κB в 87% эукариотических клеток. Напротив, инкубация с буфером приводила к транслокации менее чем в 2% HUVEC (фиг. 6A и D). Аналогичным образом, когда HUVEC обрабатывали BSA, только около 3% клеток демонстрировали транслокацию NF-κB. Интересно, что инкубация эндотелиальных клеток с растворимым белком rGroEL1 приводила к транслокации NF-κB до 92% HUVEC (рис.6Б и D). Точно так же добавление латексных шариков, покрытых rGroEL1, к HUVEC привело к транслокации фактора транскрипции примерно в 72% эукариотических клеток; Гранулы, покрытые BSA, индуцировали транслокацию только в 5% (фиг. 6C и D). Эти данные ясно указывают на то, что C. pneumoniae rGroEL1 сам по себе способен активировать эндотелиальные клетки, стимулируя транслокацию NF-κB.
РИС. 6.C. pneumoniae rGroEL1 активирует транслокацию NF-κB в HUVEC. (A) HUVEC были инфицированы C.pneumoniae или в качестве контроля с PBS в течение 2 часов. Локализацию NF-κB определяли с помощью непрямой иммунофлуоресцентной микроскопии, как описано в разделе «Материалы и методы». (B) HUVEC стимулировали 20 мкг / мл rGroEL1 или BSA в течение 5 часов. Локализация NF-κB определялась, как описано выше. (C) HUVEC стимулировали латексными шариками, покрытыми rGroEL1, или латексными шариками, покрытыми BSA, в течение 5 часов. Локализация NF-κB определялась, как описано выше. (D) Количественная оценка активации NF-κB в экспериментах, показанных на панелях A, B и C.Столбцы представляют собой отношение транслокации NF-κB в ядро к общему количеству клеток HUVEC. Каждое значение представляет собой среднее значение трех экспериментов ± стандартное отклонение.
Предварительная инкубация клеток-мишеней с C. pneumoniae белком rGroEL1 ингибирует инфекцию C. pneumoniae . Чтобы более точно охарактеризовать возможную биологическую роль подвергнутого воздействию поверхности GroEL1 во время начальной фазы инфекции, мы провели эксперименты по нейтрализации с очищенным рекомбинантный белок.Предварительная инкубация клеток HEp-2 с 20 мкг / мл rGroEL1 перед воздействием ЭБ C. pneumoniae была связана со значительным (24%) снижением количества образующихся включений по сравнению с контролем, предварительно обработанным только PBS (рис. 7А). Отрицательный контроль BSA не влиял на частоту инфицирования. Ингибирующий эффект rGroEL1 на инфекцию был дозозависимым, и максимальное ингибирование наблюдалось при концентрации rGroEL1 640 мкг / мл (фиг. 7A). Это дозозависимое снижение инфекционности предполагает, что белок GroEL1 играет роль в установлении C.pneumoniae инфекции.
РИС. 7.Растворимый хламидийный белок rGroEL1 не мешает прикреплению EB к клеткам человека, но снижает последующее инфицирование C. pneumoniae . (A) Клетки HEp-2 инкубировали с увеличивающимися количествами (от 20 мкг / мл до 640 мкг / мл) rGroEL1 в течение 2 ч до инфицирования очищенными EB C. pneumoniae . Через 60 часов после инфицирования клетки фиксировали и анализировали с помощью непрямой иммунофлуоресцентной микроскопии, как описано в разделе «Материалы и методы».Клетки HEp-2, предварительно обработанные PBS или 200 мкг / мл BSA перед инфицированием C. pneumoniae , служили контролем. Было определено количество включений хламидий, и значение, найденное для контроля PBS, было установлено на 100% ( n = 20 микроскопических полей; количество экспериментов = 4). (B) Связывание очищенных, жизнеспособных, окрашенных CFSE EB C. pneumoniae с клетками HEp-2 определяли с помощью проточного цитометрического анализа. В контрольных экспериментах отслеживали прикрепление EB C. pneumoniae к клеткам HEp-2, инкубированным с PBS, 200 мкг / мл БСА или 500 мкг / мл гепарина, и сравнивали со скоростью связывания в образцах, в которых 20 мкг / мл или Добавляли 200 мкг / мл белка rGroEL1 (количество экспериментов = 4).Планки погрешностей указывают стандартные отклонения.
Предварительная инкубация клеток-мишеней с белком rGroEL1 не мешает адгезии EB C. pneumoniae . Снижение инфекционности, опосредованное rGroEL1, может быть достигнуто двумя способами, которые не исключают друг друга: добавление растворимого rGroEL1 также может напрямую блокируют рецепторы на поверхности клеток HEp-2, которые необходимы для прикрепления и / или интернализации EB, или могут вызвать преждевременную активацию путей, которые отрицательно влияют на установление инфекции (например,g., через активацию NF-κB, наблюдаемую в HUVEC [Рис. 6]). Чтобы проверить, является ли локализованный на поверхности хламидийный GroEL1 необходимым условием для адгезии EB к клеткам-хозяевам, были выполнены проточно-цитометрические анализы связывания жизнеспособных EB, меченных CFSE, с клетками HEp-2. Включение гепарина привело к 95% уменьшению прикрепления EB, тогда как добавление rGroEL1 в концентрации 20 мкг / мл или 200 мкг / мл не имело ингибирующего эффекта на связывание EB с клетками Hep-2 (фиг. 7B). Этот результат указывает на то, что GroEL1, по-видимому, не играет роли в прикреплении EB.
ОБСУЖДЕНИЕ
Это исследование демонстрирует, что C. pneumoniae GroEL1 связан с поверхностью инфекционных EB и может быть обнаружен во включении на более позднем этапе развития инфекции. Эксперименты по фракционированию показали, что примерно половина общего GroEL1, обнаруженного в EB, локализована на их поверхности. Латексные шарики, покрытые rGroEL1, прикреплялись к клеткам HEp-2 дозозависимым образом, в то время как шарики обрабатывались двумя другими гомологами GroEL из C.pneumoniae и GroEL из E. coli не смогли этого сделать. Связывающие свойства GroEL1 были подтверждены с использованием тестов дрожжевого дисплея. Инкубация HUVEC с растворимым rGroEL1 или с шариками, покрытыми rGroEL1, приводила к транслокации NF-κB в ядро. Наконец, инфицирование C. pneumoniae подавлялось дозозависимым образом путем предварительной инкубации клеток HEp-2 с растворимым белком rGroEL1. Однако прямые эксперименты по адгезии показали, что связывание EB с клетками HEp-2, по-видимому, не опосредуется GroEL1.
Белок GroEL1 связан с поверхностью EB и хламидийным включением во время цикла развития. Наши результаты подтверждают и расширяют ранее полученные данные о локализации GroEL1. Ранние биохимические эксперименты уже предполагали, что GroEL1 связан с фракциями внешней мембраны инфекционных EB из C. trachomatis и C. psittaci , из которых он может быть высвобожден с помощью мягких детергентов (3). Совсем недавно исследования с помощью электронной микроскопии обнаружили C.trachomatis GroEL1 во включениях не только в цитоплазме бактерий, но и в хламидийной оболочке и в просвете включения (61). Это недавно было описано также для C. pneumoniae GroEL1. В этом случае белок был обнаружен с помощью электронной микроскопии во включениях репликационно-пермиссивных эпителиальных клеток HEp-2, а также во внеклеточном пространстве (12). Наши данные иммунолокализации также доказывают, что белок C. pneumoniae GroEL1 является включением, но вне хламидийных частиц, а также на поверхности инфекционных EB.Кроме того, наши данные свидетельствуют о том, что по крайней мере 50% GroEL1 в изолированных инфекционных EB связаны с поверхностью, поскольку его можно восстановить, промывая EB PBS. Лизис бактериальных клеток был исключен как возможный источник этой фракции белка, поскольку было обнаружено, что внутрихламидийный белок, DnaK, не был обогащен растворимой фазой в этих условиях. Наиболее вероятно, что относительное количество GroEL1, связанного с поверхностью EB, выше in vivo, поскольку EB, используемые в экспериментах по фракционированию, были очищены с использованием градиентов Перколла, которые могли удалить часть GroEL1 с их поверхностей.Фракция GroEL1, которая остается связанной с EB, предположительно находится в цитоплазме бактерий, где она, вероятно, служит молекулярным шаперонином. Тот факт, что хламидийный GroEL1 (но не GroEL2 и GroEL3) может функционально заменять чувствительный к температуре эндогенный GroEL в мутанте E. coli , убедительно свидетельствует о том, что он может функционировать как шаперонин (37). Следовательно, дифференциальная компартментализация GroEL1 д. Предполагать двойную роль этого белка, и, таким образом, GroEL1 может принадлежать к растущей группе «подрабатывающих белков» (34).
Внеклеточная локализация и роль GroEL1 в молекулярном перекрестном взаимодействии между патогенным микроорганизмом и его клеткой-хозяином являются необычными особенностями для белка теплового шока, но в настоящее время существует несколько прецедентов такой многофункциональности. Таким образом, внеклеточная локализация GroEL была описана в Bartonella bacilliformis , Helicobacter pylori , Legionella pneumophila и Francisella tularensis (см. Ссылки 21, 27, 29, 48 и 54). H. pylori , Clostridium difficile , Haemophilus ducreyi и Salmonella enterica серовар Typhimurium все используют поверхностные GroEL-подобные белки, чтобы способствовать прикреплению к клеткам человека (см. Ссылку 27). У молочнокислой бактерии Lactobacillus johnsonii , естественного обитателя желудочно-кишечного тракта млекопитающих, белок GroEL был обнаружен на бактериальной поверхности, где он может способствовать прикреплению к слизи и эпителиальным клеткам, стимулировать секрецию цитокинов и опосредовать агрегацию желудочного сока. патоген H.pylori (4). Аналогичным образом GroEL-подобный белок, обнаруженный на поверхности вирулентных штаммов L. pneumophila , действует как адгезин и опосредует интернализацию и специфический перенос латексных частиц в клетки HeLa (21). У эукариотического диморфного грибкового патогена Histoplasma capsulatum локализованный на поверхности GroEL-подобный белок опосредует прикрепление факультативно внутриклеточного гриба к β 2 рецепторам интегрина (CD11 / CD18) на макрофагах (49). Таким образом, возможно, что у различных, в основном патогенных, организмов GroEL и GroEL-подобные белки были диверсифицированы в ходе эволюции и развили новые функции, связанные с различными паттернами субклеточной локализации.Например, в эндосимбионте Enterobacter aerogenes GroEL-подобный белок отличается в 11 положениях от E. coli GroEL и обнаруживается как внутри-, так и внеклеточно, действуя как парализующий токсин на насекомых (87).
Белки GroEL, которые считаются прототипами цитозольных белков, вероятно, не единственные цитозольные белки, которые также могут направляться на поверхность бактериальной клетки. Недавно было обнаружено, что фактор удлинения трансляции EF-Tu присутствует на поверхности л.johnsonii , опосредуя их прикрепление к клеткам человека (24). Характеристика субпротеома клеточной стенки Listeria monocytogenes выявила присутствие GroEL, енолазы и EF-Tu среди других типичных «цитоплазматических» белков (68). Как и C. pneumoniae, GroEL1, эти белки не несут никаких очевидных сигналов секреции или известных поверхностно-связывающих доменов. Таким образом, механизм, с помощью которого они достигают поверхности бактерий, остается неясным. Было высказано предположение, однако, что эти «подрабатывающие» белки могут экспортироваться транспортерами ABC или путем пассивного высвобождения при осмотическом стрессе (68).Альтернативно, эти белки могут экспортироваться с помощью так называемого пути секреции типа VI, описанного для грамотрицательных бактерий. Многие из них высвобождают везикулы из своей внешней мембраны в качестве секреторных носителей для компонентов, имеющих отношение к бактериальной адгезии и инвазии, цитотоксичности и ответам хозяина (45).
rGroEL1 активирует первичные эндотелиальные клетки человека. В настоящем исследовании мы продемонстрировали, что C. pneumoniae, rGroEL1 опосредует активацию NF-κB в HUVEC, и что растворимый GroEL1 и латексные шарики, покрытые белком, почти одинаково эффективны при этом. .Активация и транслокация NF-κB в ядро эукариотических клеток, инфицированных C. pneumoniae , были продемонстрированы в различных исследованиях (6, 40, 44). Активация NF-κB является важным этапом в эукариотической передаче сигналов, вызванной патогенами, наблюдаемой для различных патогенов, таких как H. pylori , энтерогеморрагическая E. coli или Campylobacter jejuni (38, 53, 67) .
Активация NF-κB во время инфекции C. pneumoniae была описана для множества различных типов клеток (например,g., эпителиальные клетки, HUVEC, человеческие моноциты и клетки Mono Mac) (22, 44, 82, 83). Активация NF-κB в HUVEC происходит очень быстро, так как ее можно было обнаружить в течение 15 минут после заражения C. pneumoniae (44). Точно так же в клетках Mono Mac активация NF-κB достигла пика в течение первых 60 мин после заражения (83, 82). Это убедительно свидетельствует о том, что внутриклеточные сигнальные события, ведущие к транслокации NF-κB, индуцируются при начальном контакте хламидийных частиц с эукариотическими клетками.В этом отношении активация NF-κB латексными шариками, покрытыми GroEL1, может имитировать контакт EB C. pneumoniae с эукариотическими клетками.
Инфекция C. pneumoniae запускает дополнительные ответы клетки-хозяина, такие как фосфорилирование митоген-активируемой киназы p40 / 42; секреция интерлейкина-1 (ИЛ-1), ИЛ-8, ИЛ-10 и хемоаттрактантного белка моноцитов 1; и экспрессия рецепторов клеточной поверхности, таких как ICAM, VCAM, E-селектин и тканевый фактор (6, 9, 15, 41, 44, 56, 66, 83).Некоторые из этих ответов хозяйских клеток также наблюдались при инкубации эукариотических клеток с C. trachomatis GroEL1 (41, 66). Это предполагает, что такая активация может быть общим свойством хламидийных белков GroEL1. Действительно, C. pneumoniae rGroEL1 инициирует секрецию IL-12 и фактора некроза опухоли через TLR в дендритных клетках и активирует эндотелиальные клетки сосудов для секреции цитокинов IL-1β, IL-6 и IL-8 и экспрессии белков адгезии на клеточной поверхности. (12, 51).Следовательно, возможной моделью является такая, в которой поверхностная локализация C. pneumoniae GroEL1 способствует вызванной патогеном активации эукариотических клеток-хозяев при прикреплении хламидий и впоследствии запускает пути внутриклеточной передачи сигнала внутри эукариотических клеток, как описано другими ( 31, 40, 44, 56, 66, 82, 83). Природа рецептора GroEL1 на клетке-хозяине до сих пор неясна. Однако сообщалось, что GroEL1 из C. pneumoniae связывается с CD14, TLR4 и / или TLR2 (9, 12, 41, 51).
Поверхностная локализация C. pneumoniae GroEL1 на EB и активация HUVEC с помощью rGroEL1 убедительно свидетельствуют о том, что GroEL1 играет важную роль в начальной фазе взаимодействия патоген-хозяин, и наши данные прямо подтверждают эту идею, поскольку предварительная обработка клеток HEp-2 rGroEL1 значительно снижает последующую инфекционность. Однако предварительная обработка эпителиальных клеток rGroEL1 не ингибировала адгезию инфекционных EB, что позволяет предположить, что GroEL1, локализованный на поверхности бактериальных клеток, не требуется для прикрепления EB к клеткам HEp-2.Однако можно предположить, что связывание GroEL1 с эукариотическими рецепторами может оказывать влияние на более поздние стадии становления инфекции; например, белок может иметь отношение к интернализации EB или модификации раннего включения. Таким образом, преждевременный и / или гиперактивный ответ, индуцированный в клетках HEp-2 с помощью rGroEL1 до добавления EB, вполне может быть вредным для хозяина и / или для успешного инфицирования. Тот факт, что rGroEL1 может активировать транслокацию NF-κB в HUVEC с кинетикой, аналогичной кинетике, обнаруженной при нормальном C.pneumoniae подтверждает эту сигнальную гипотезу.
В заключение, наши данные свидетельствуют о том, что фракция белка C. pneumoniae GroEL1 локализована на поверхности бактериальной клетки и играет важную роль в возникновении хламидийной инфекции, возможно, путем индукции ранних сигнальных событий, которые помогают преобразовать эукариотическую клетку в гостеприимную среду, подходящую для успешного заражения.
БЛАГОДАРНОСТИ
Эта работа была поддержана Deutsche Forschungsgemeinschaft, Sonderforschungsbereich 590, проект номер C5 (J.H.H.) и сеть Era-NET PathoGenoMics ECIBUG.
Благодарим Урсулу Флейг за критическое прочтение рукописи. Мы очень ценим помощь Александры Кульяк в создании плазмиды pAC-2.
У авторов нет коммерческих или иных объединений, которые могли бы представлять конфликт интересов.
СНОСКИ
- Получено 9 октября 2007 г.
- Принято 24 февраля 2008 г.
- Авторское право © Американское общество микробиологии, 2008 г.
ССЫЛКИ
- 1.↵
Болдуин, А.С., младший 2001. Введение в серию: фактор транскрипции NF-kappaB и болезнь человека. J. Clin. Расследование. 107 : 3-6.
- 2.↵
Баттейгер, Б. Э., В. Дж. Т. Ньюхолл и Р. Б. Джонс. 1985. Различия в белках внешней мембраны венерической лимфогранулемы и биоваров трахомы Chlamydia trachomatis . Заразить. Иммун. 50 : 488-494.
- 3.↵
Бавойл, П., Р. С. Стивенс и С. Фалькоу. 1990. Растворимый 60 килодальтонный антиген Chlamydia spp. является гомологом Escherichia coli GroEL. Мол. Microbiol. 4 : 461-469.
- 4.↵
Бергонцелли, Г. Э., Д. Гранато, Р. Д. Придмор, Л. Ф. Марвин-Гай, Д. Донникола и И. Э. Кортези-Тейлаз. 2006. GroEL Lactobacillus johnsonii La1 (NCC 533) связан с клеточной поверхностью: потенциальная роль во взаимодействиях с хозяином и желудочным патогеном Helicobacter pylori .Заразить. Иммун. 74 : 425-434.
- 5.↵
Birkelund, S., A. G. Lundemose и G. Christiansen. 1990. 75-килодальтонный цитоплазматический полипептид L2 Chlamydia trachomatis представляет собой DnaK-подобный белок. Заразить. Иммун. 58 : 2098-2104.
- 6.↵
Блессинг, Э., К. К. Куо, Т. М. Линь, Л. А. Кэмпбелл, Ф. Би, Б. Чезебро и М. Э. Розенфельд. 2002. Образование пенистых клеток подавляет рост Chlamydia pneumoniae, но не снижает индуцированную Chlamydia pneumoniae секрецию провоспалительных цитокинов.Тираж 105 : 1976–1982 годы.
- 7.↵
Brunham, R.C. и R.W. Peeling. 1994. Антигены Chlamydia trachomatis: роль в иммунитете и патогенезе. Заразить. Агенты Дис. 3 : 218-233.
- 8.↵
Bukau, B., and A. L. Horwich. 1998. Сопровождающие аппараты Hsp70 и Hsp60. Клетка 92 : 351-366.
- 9.↵
Булут Ю., Фор Э., Томас Л., Х.Карахаши, К. С. Мичельсен, О. Эквилс, С. Г. Моррисон, Р. П. Моррисон и М. Ардити. 2002. Хламидийный белок теплового шока 60 активирует макрофаги и эндотелиальные клетки через Toll-подобный рецептор 4 и MD2 по MyD88-зависимому пути. J. Immunol. 168 : 1435-1440.
- 10.↵
Кэмпбелл, Л. А., и К. К. Куо. 2006. Взаимодействие Chlamydia с клетками-хозяевами, которые опосредуют прикрепление и захват, с. 505-522. В п.М. Бавойл и П. Б. Уайрик (ред.), Геномика и патогенез хламидий. Horizon Bioscience, Норфолк, Соединенное Королевство.
- 11.↵
Конант, К. Г. и Р. С. Стивенс. 2007. Для прикрепления хламидий к клеткам млекопитающих требуется протеин-дисульфид-изомераза. Клетка. Microbiol. 9 : 222-232.
- 12.↵
Коста, К. П., К. Дж. Киршнинг, Д. Буш, С. Дурр, Л. Дженнен, У. Хайнцманн, С. Пребек, Х. Вагнер и Т. Митке. 2002. Роль хламидийного белка теплового шока 60 в стимуляции клеток врожденного иммунитета Chlamydia pneumoniae. Евро. J. Immunol. 32 : 2460-2470.
- 13.↵
Крейн, Д. Д., Дж. Х. Карлсон, Э. Р. Фишер, П. Бавойл, Р. К. Сиа, К. Тан, К. К. Куо и Г. Д. Колдуэлл. 2006. Полиморфный мембранный белок D Chlamydia trachomatis представляет собой обще-нейтрализующий антиген. Proc. Natl. Акад. Sci. США 103 : 1894-1899 гг.
- 14.↵
Дэвис, К. Х., Дж. Э. Раулстон и П. Б. Уайрик. 2002. Протеин-дисульфидизомераза, компонент рецепторного комплекса эстрогена, связана с сероваром E Chlamydia trachomatis, , прикрепленным к эпителиальным клеткам эндометрия человека. Заразить. Иммун. 70 : 3413-3418.
- 15.↵
Dechend, R., M. Maass, J. Gieffers, R. Dietz, C. Scheidereit, A. Leutz и D.C. Gulba. 1999. Инфекция Chlamydia pneumoniae гладких мышц сосудов и эндотелиальных клеток активирует NF-kappaB и индуцирует тканевый фактор и экспрессию PAI-1: потенциальная связь с ускоренным атеросклерозом.Тираж 100 : 1369-1373.
- 16.↵
Dersch, P., and R.R. Isberg. 1999. Область белка инвазина Yersinia pseudotuberculosis усиливает опосредованное интегрином поглощение клетками млекопитающих и способствует самоассоциации. EMBO J. 18 : 1199-1213.
- 17.↵
Eckert, L.O., S.E. Hawes, P. Wolner Hanssen, D.M. Money, R.W. Peeling, R.C. Brunham, C.E.Stevens, D.A. Eschenbach, W.Э. Штамм. 1997. Распространенность и корреляты антител к хламидийному белку теплового шока у женщин, посещающих клиники по лечению заболеваний, передающихся половым путем, и женщин с подтвержденным воспалительным заболеванием органов малого таза. J. Infect. Дис. 175 : 1453-1458.
- 18.↵
Entian, KD, T. Schuster, JH Hegemann, D. Becher, H. Feldmann, U. Guldener, R. Gotz, M. Hansen, CP Hollenberg, G. Jansen, W. Крамер, С. Кляйн, П. Коттер, Дж. Крике, Х. Лаунхардт, Г. Маннхаупт, А.Майерл, П. Мейер, В. Мьюз, Т. Мундер, Р. К. Ниденталь, М. Рамезани Рад, А. Ромер, А. Ромер, А. Хиннен и др. 1999. Функциональный анализ 150 делеционных мутантов в Saccharomyces cerevisiae с помощью системного подхода. Мол. Genet Genet. 262 : 683-702.
- 19.↵
Fadel, S., and A. Eley. 2007. Белок OmcB Chlamydia trachomatis представляет собой гликозаминогликан-зависимый адгезин, доступный на поверхности. J. Med. Microbiol. 56 : 15-22.
- 20.№
Флешнер М. и Дж. Д. Джонсон. 2005. Эндогенный внеклеточный белок теплового шока 72: высвобождающий сигнал (ы) и функция. Int. J. Гипертермия 21 год : 457-471.
- 21.↵
Garduno, R. A., E. Garduno и P. S. Hoffman. 1998. Поверхностно-связанный шаперонин hsp60 Legionella pneumophila опосредует инвазию в модели клеток HeLa. Заразить. Иммун. 66 : 4602-4610.
- 22.№
Gencay, M. M., M. Tamm, A. Glanville, A. P. Perruchoud и M. Roth. 2003. Chlamydia pneumoniae активирует пролиферацию эпителиальных клеток через NF-κB и рецептор глюкокортикоидов. Заразить. Иммун. 71 : 5814-5822.
- 23. ↵
Джерард, Х. К., К. Л. Вильдт, Дж. А. Уиттум-Хадсон, З. Лай, Дж. Агер и А. П. Хадсон. 2005. Нагрузка Chlamydia pneumoniae в головном мозге Альцгеймера варьируется в зависимости от генотипа APOE.Microb. Патог. 39 : 19-26.
- 24.↵
Гранато Д., Г. Э. Бергонцелли, Р. Д. Придмор, Л. Марвин, М. Руве и И. Э. Кортези-Тейлаз. 2004. Связанный с клеточной поверхностью фактор удлинения Tu опосредует прикрепление Lactobacillus johnsonii NCC533 (La1) к клеткам кишечника и муцинам человека. Заразить. Иммун. 72 : 2160-2169.
- 25.↵
Grayston, J. T. 1989. Chlamydia pneumoniae, штамм TWAR.Грудь 95 : 664-669.
- 26.↵
Хан, Д. Л. и Р. Макдональд. 1998. Может ли острая инфекция дыхательных путей Chlamydia pneumoniae вызвать хроническую астму? Аня. Allergy Asthma Immunol. 81 год : 339-344.
- 27.↵
Хендерсон Б., Э. Аллан и А. Р. Коутс. 2006. Стрессовые войны: прямая роль хозяина и бактериальных молекулярных шаперонов в бактериальной инфекции. Заразить. Иммун. 74 : 3693-3706.
- 28.↵
Хорнер, П. Дж., Д. Каин, М. МакКлюр, Б. Дж. Томас, К. Гилрой, М. Али, Дж. Н. Вебер и Д. Тейлор-Робинсон. 1997. Ассоциация антител к белку теплового шока Chlamydia trachomatis 60 кДа с хроническим негонококковым уретритом. Clin. Заразить. Дис. 24 : 653-660.
- 29.↵
Уэска, М., С. Борджиа, П. Хоффман и К. А. Лингвуд. 1996. Кислый pH изменяет специфичность связывания рецептора Helicobacter pylori : бинарная модель адгезии, в которой поверхностные белки теплового шока (стресса) опосредуют распознавание сульфатидов при колонизации желудка.Заразить. Иммун. 64 : 2643-2648.
- 30.↵
Hybiske, K., and R. S. Stephens. 2007. Механизмы выхода из клетки-хозяина внутриклеточной бактерией Chlamydia. Proc. Natl. Акад. Sci. США 104 : 11430-11435.
- 31.↵
Jahn, H.U., M. Krull, F. N. Wuppermann, A.C. Klucken, S. Rosseau, J. Seybold, J. H. Hegemann, C.A. Jantos, and N. Suttorp. 2000. Инфекция и активация эпителиальных клеток дыхательных путей Chlamydia pneumoniae.J. Infect. Дис. 182 : 1678–1687.
- 32.↵
Джантос, К. А., С. Хек, Р. Роггендорф, М. Сен-Гупта и Дж. Х. Хегеманн. 1997. Антигенный и молекулярный анализ различных штаммов Chlamydia pneumoniae . J. Clin. Microbiol. 35 год : 620-623.
- 33.↵
Jantos, C. A., C. Krombach, F. N. Wuppermann, A. Gardemann, S. Bepler, H. Asslan, J. H. Hegemann, and W. Haberbosch. 2000 г.Антительный ответ на белок теплового шока 60 кДа Chlamydia pneumoniae у пациентов с ишемической болезнью сердца. J. Infect. Дис. 181 : 1700–1705.
- 34.↵
Джеффри, К. Дж. 1999. Подработка белков. Trends Biochem. Sci. 24 : 8-11.
- 35.↵
Kaiser, C., S. Michaelis, and A. Mitchell. 1994. Методы в генетике дрожжей. Лабораторная пресса Колд-Спринг-Харбор, Колд-Спринг-Харбор, Нью-Йорк.
- 36.
Kalman, S., W. Mitchell, R. Marathe, C. Lammel, J. Fan, R. W. Hyman, L. Olinger, J. Grimwood, R. W. Davis и R. S. Stephens. 1999. Сравнительные геномы Chlamydia pneumoniae и C. trachomatis. Nat. Genet. 21 год : 385-389.
- 37.↵
Карунакаран, К. П., Я. Ногучи, Т. Д. Рид, А. Черкасов, Дж. Кви, К. Шен, К. К. Нельсон и Р. К. Брунхэм. 2003. Молекулярный анализ множественных белков GroEL хламидий.J. Bacteriol. 185 : 1958-1966 гг.
- 38.↵
Кейтс, С., Ю. С. Хитти, М. Аптон и К. П. Келли. 1997. Инфекция Helicobacter pylori активирует NF-каппа B в эпителиальных клетках желудка. Гастроэнтерология 113 : 1099-1109.
- 39.↵
Knudsen, K., A. S. Madsen, P. Mygind, G. Christiansen и S. Birkelund. 1999. Идентификация двух новых генов, кодирующих белки внешней мембраны от 97 до 99 килодальтон Chlamydia pneumoniae .Заразить. Иммун. 67 : 375-383.
- 40.↵
Кол, А., Т. Бурсье, А. Х. Лихтман и П. Либби. 1999. Хламидийный белок 60-х годов теплового шока человека активирует эндотелий сосудов человека, гладкомышечные клетки и макрофаги. J. Clin. Расследование. 103 : 571-577.
- 41.↵
Кол, А., А. Х. Лихтман, Р. В. Финберг, П. Либби и Э. А. Курт-Джонс. 2000. Белок теплового шока (HSP) 60 активирует врожденный иммунный ответ: CD14 является важным рецептором для активации HSP60 мононуклеарных клеток.J. Immunol. 164 : 13-17.
- 42.↵
Корппи, М., Т. Хейсканен Косма, Э. Ялонен, П. Сайкку, М. Лейнонен, П. Халонен и П. Х. Макела. 1993. Этиология внебольничной пневмонии у детей, находящихся на стационарном лечении. Евро. J. Pediatr. 152 : 24-30.
- 43.↵
Krull, M., P. Bockstaller, FN Wuppermann, AC Klucken, J. Muhling, B. Schmeck, J. Seybold, C. Walter, M. Maass, S. Rosseau, J .Х. Хегеманн, Н. Сутторп и С. Хиппенштиль. 2006. Механизмы опосредованного Chlamydophila pneumoniae высвобождения GM-CSF в эпителиальных клетках бронхов человека. Являюсь. J. Respir. Cell Mol. Биол. 34 : 375-382.
- 44.↵
Krüll, M., A.C. Klucken, F. N. Wuppermann, O. Fuhrmann, C.Magerl, J. Seybold, S. Hippenstiel, J. H. Hegemann, C. A. Jantos и N. Suttorp. 1999. Пути передачи сигнала, активируемые в эндотелиальных клетках после заражения Chlamydia pneumoniae.J. Immunol. 162 : 4834-4841.
- 45.↵
Kuehn, M. J., and N.C. Kesty. 2005. Бактериальные везикулы наружной мембраны и взаимодействие хозяина-патогена. Genes Dev. 19 : 2645-2655.
- 46.↵
Куо, К. К., Л. А. Джексон, Л. А. Кэмпбелл и Дж. Т. Грейстон. 1995. Chlamydia pneumoniae (TWAR). Clin. Microbiol. Ред. 8 : 451-461.
- 47.↵
Куо, К.К., Н. Такахаши, А. Ф. Свонсон, Ю. Озеки и С. И. Хакомори. 1996. N-связанный олигосахарид высокоманнозного типа, экспрессируемый в основном белке внешней мембраны Chlamydia trachomatis, опосредует прикрепление и инфекционность микроорганизма к клеткам Hela. J. Clin. Расследование. 98 : 2813-2818.
- 48.↵
Ли Б. Ю., М. А. Хорвиц и Д. Л. Клеменс. 2006. Идентификация, рекомбинантная экспрессия, иммунолокализация в макрофагах и Т-клеточная чувствительность основных внеклеточных белков Francisella tularensis .Заразить. Иммун. 74 : 4002-4013.
- 49.
Лонг, К. Х., Ф. Дж. Гомес, Р. Э. Моррис и С. Л. Ньюман. 2003. Идентификация белка теплового шока 60 как лиганда Histoplasma capsulatum, который опосредует связывание с рецепторами CD18 на макрофагах человека. J. Immunol. 170 : 487-494.
- 50.↵
Lundemose, A. G., S. Birkelund, P. M. Larsen, S. J. Fey и G. Christiansen. 1990. Характеристика и идентификация ранних белков в сероваре L2 Chlamydia trachomatis с помощью двумерного гель-электрофореза.Заразить. Иммун. 58 : 2478-2486.
- 51.↵
Магуайр, М., С. Пул, А. Р. Коутс, П. Тормей, К. Уиллер-Джонс и Б. Хендерсон. 2005. Сравнительная клеточная сигнальная активность ультрачистых рекомбинантных белков шаперонина 60 от прокариот и эукариот. Иммунология 115 : 231-238.
- 52.↵
Melby, K. K., T. K. Kvien, A. Glennas, and G. Anestad. 1999. Chlamydia pneumoniae как триггер реактивного артрита.Сканд. J. Infect. Дис. 31 год : 327-328.
- 53.↵
Меллитс, К. Х., Дж. Маллен, М. Ванд, Г. Армбрустер, А. Патель, П. Л. Коннертон, М. Скелли и И. Ф. Коннертон. 2002. Активация фактора транскрипции NF-kappaB с помощью Campylobacter jejuni. Микробиология 148 : 2753-2763.
- 54.↵
Минник, М. Ф., Л. С. Смитерман и Д. С. Самуэльс. 2003. Митогенный эффект Bartonella bacilliformis на эндотелиальные клетки сосудов человека и участие GroEL.Заразить. Иммун. 71 : 6933-6942.
- 55.↵
Moelleken, K., and J. H. Hegemann. 2008. Белок внешней мембраны Chlamydia OmcB необходим для адгезии и проявляет биовар-специфические различия в связывании гликозаминогликанов. Мол. Microbiol. 67 : 403-419.
- 56.
Molestina, R. E., R. D. Miller, J. A. Ramirez и J. T. Summersgill. 1999. Инфекция эндотелиальных клеток человека Chlamydia pneumoniae стимулирует трансэндотелиальную миграцию нейтрофилов и моноцитов.Заразить. Иммун. 67 : 1323-1330.
- 57.↵
Моррисон, Р. П. 1991. Хламидийный hsp60 и иммунопатогенез хламидийной болезни. Семин. Иммунол. 3 : 25-33.
- 58.↵
Pfister, G., C. M. Stroh, H. Perschinka, M. Kind, M. Knoflach, P. Hinterdorfer и G. Wick. 2005. Обнаружение HSP60 на поверхности мембраны стрессированных эндотелиальных клеток человека с помощью атомной силы и конфокальной микроскопии.J. Cell Sci. 118 : 1587–1594.
- 59.↵
Рэнк, Р. Г., К. Дашер, А. К. Боулин и П. М. Бавойл. 1995. Системная иммунизация Hsp60 изменяет развитие хламидийной глазной болезни. Расследование. Офтальмол. Vis. Sci. 36 : 1344-1351.
- 60.
Raulston, J. E., C. H. Davis, T. R. Paul, J. D. Hobbs и P. B. Wyrick. 2002. Доступность поверхности 70-килодальтонного белка теплового шока Chlamydia trachomatis после восстановления дисульфидных связей белка внешней мембраны.Заразить. Иммун. 70 : 535-543.
- 61.↵
Raulston, J. E., T. R. Paul, S. T. Knight и P. B. Wyrick. 1998. Локализация Chlamydia trachomatis белков теплового шока 60 и 70 во время инфицирования линии эпителиальных клеток эндометрия человека in vitro. Заразить. Иммун. 66 : 2323-2329.
- 62.↵
Рид, Т. Д., Р. К. Брунхэм, К. Шен, С. Р. Гилл, Дж. Ф. Гейдельберг, О. Уайт, Э. К.Хики, Дж. Петерсон, Т. Аттербэк, К. Берри, С. Басс, К. Линхер, Дж. Вайдман, Х. Кури, Б. Крейвен, К. Боуман, Р. Додсон, М. Гвинн, В. Нельсон, Р. ДеБой, Дж. Колоней, Дж. Макларти, С.Л. Зальцберг, Дж. Эйзен и С.М. Фрейзер. 2000. Последовательности генома Chlamydia trachomatis MoPn и Chlamydia pneumoniae AR39. Nucleic Acids Res. 28 год : 1397-1406.
- 63.↵
Reynolds, D. J., and J. H. Pearce. 1991. Эндоцитарные механизмы, используемые хламидиями, и их влияние на индукцию продуктивной инфекции.Заразить. Иммун. 59 : 3033-3039.
- 64.↵
Сайкку П., М. Лейнонен, К. Маттила, М. Р. Экман, М. С. Ниеминен, П. Х. Макела, Дж. К. Хуттунен и В. Валтонен. 1988. Серологические доказательства связи нового хламидиоза, TWAR, с хронической ишемической болезнью сердца и острым инфарктом миокарда. Ланцет II : 983-986.
- 65.↵
Санчес-Кампильо, М., Л. Бини, М. Командуччи, Р. Раггиаши, Б.Марзокки, В. Паллини и Г. Ратти. 1999. Идентификация иммунореактивных белков Chlamydia trachomatis с помощью вестерн-блоттинга двухмерной карты электрофореза с сыворотками пациентов. Электрофорез 20 : 2269-2279.
- 66.↵
Сасу, С., Д. Ла Верда, Н. Куреши, Д. Т. Голенбок и Д. Бизли. 2001. Chlamydia pneumoniae и хламидийный белок теплового шока 60 стимулируют пролиферацию гладкомышечных клеток сосудов человека через Toll-подобный рецептор 4 и активацию митоген-активируемой протеинкиназы p44 / p42.Circ. Res. 89 : 244-250.
- 67.↵
Savkovic, S. D., A. Koutsouris, and G. Hecht. 1997. Активация NF-kappaB в эпителиальных клетках кишечника энтеропатогенной Escherichia coli. Являюсь. J. Physiol. 273C : 1160-1167.
- 68.↵
Schaumburg, J., O. Diekmann, P. Hagendorff, S. Bergmann, M. Rohde, S. Hammerschmidt, L. Jansch, J. Wehland и U. Karst. 2004. Субпротеом клеточной стенки Listeria monocytogenes.Протеомика 4 : 2991-3006.
- 69.↵
Schiestl, R.H. и R.D. Gietz. 1989. Высокоэффективная трансформация интактных дрожжевых клеток с использованием одноцепочечных нуклеиновых кислот в качестве носителя. Curr. Genet. 16 : 339-346.
- 70.↵
Schmiel, D. H., S. T. Knight, J. E. Raulston, J. Choong, C.H. Davis и P. B. Wyrick. 1991. Рекомбинантные клоны Escherichia coli , экспрессирующие продукты гена Chlamydia trachomatis , прикрепляются к эпителиальным клеткам эндометрия человека.Заразить. Иммун. 59 : 4001-4012.
- 71.↵
Schnitger, K., F. Njau, U. Wittkop, A. Liese, J. G. Kuipers, A. Thiel, M. A. Morgan, H. Zeidler, and A. D. Wagner. 2007. Окрашивание элементарных тел Chlamydia trachomatis: подходящий метод для идентификации инфицированных моноцитов человека с помощью проточной цитометрии. J. Microbiol. Методы 69 : 116-121.
- 72.↵
Шерман, Ф. 1991. Начало работы с дрожжами.Методы Энзимол. 194 : 3-21.
- 73.↵
Шин, Б.К., Х. Ван, А.М. Йим, Ф. Ле Наур, Ф. Брихори, Дж. Х. Янг, Р. Чжао, Э. Пуравс, Дж. Тра, К. У. Майкл, Д. Мисек , и С.М. Ханаш. 2003. Глобальное профилирование протеома клеточной поверхности раковых клеток обнаруживает большое количество белков с функцией шаперона. J. Biol. Chem. 278 : 7607-7616.
- 74.↵
Sikorski, R. S., and P. Hieter. 1989. Система челночных векторов и штаммов-хозяев дрожжей, разработанная для эффективного манипулирования ДНК в Saccharomyces cerevisiae. Генетика 122 : 19-27.
- 75.↵
Stephens, RS, S. Kalman, C. Lammel, J. Fan, R. Marathe, L. Aravind, W. Mitchell, L. Olinger, RL Tatusov, Q. Zhao, EV Кунин и Р.В. Дэвис. 1998. Последовательность генома облигатного внутриклеточного патогена человека: Chlamydia trachomatis . Наука 282 : 754-759.
- 76.↵
Стивенс Р. С., К. Кошияма, Э. Льюис и А. Кубо. 2001. Гепарин-связывающий белок внешней мембраны хламидий. Мол. Microbiol. 40 : 691-699.
- 77.↵
Stratton, C. W., and S. Sriram. 2003. Связь Chlamydia pneumoniae с заболеванием центральной нервной системы. Микробы заражают. 5 : 1249-1253.
- 78.↵
Су, Х., Л. Раймонд, Д.Д. Роки, Э. Фишер, Т. Хакштадт и Х. Д. Колдуэлл. 1996. Рекомбинантный белок главной внешней мембраны Chlamydia trachomatis связывается с рецепторами гепарансульфата на эпителиальных клетках. Proc. Natl. Акад. Sci. США 93 : 11143-11148.
- 79. №
Том Д. Х., Дж. Т. Грейстон, Л. А. Кэмпбелл, К. К. Куо, В. К. Диван и С. П. Ван. 1994. Респираторная инфекция Chlamydia pneumoniae у взрослых амбулаторных пациентов среднего и старшего возраста. Евро. J. Clin.Microbiol. Заразить. Дис. 13 : 785-792.
- 80.↵
Ting, L. M., R. C. Hsia, C. G. Haidaris и P. M. Bavoil. 1995. Взаимодействие белков внешней оболочки GPIC Chlamydia psittaci с поверхностью клеток HeLa. Заразить. Иммун. 63 : 3600-3608.
- 81.↵
Вабулас, Р. М., П. Ахмад-Нежад, К. да Коста, Т. Митке, К. Дж. Киршнинг, Х. Хакер и Х. Вагнер. 2001. Эндоцитозированные HSP60 используют Toll-подобный рецептор 2 (TLR2) и TLR4 для активации сигнального пути рецептора Toll / интерлейкина-1 в клетках врожденного иммунитета.J. Biol. Chem. 276 : 31332-31339.
- 82.↵
Wahl, C., S. Maier, R. Marre и A. Essig. 2003. Chlamydia pneumoniae индуцирует экспрессию ингибитора апоптоза 2 (c-IAP2) в линии моноцитарных клеток человека посредством NF-kappaB-зависимого пути. Int. J. Med. Microbiol. 293 : 377-381.
- 83.↵
Wahl, C., F. Oswald, U. Simnacher, S. Weiss, R. Marre и A. Essig. 2001. Выживаемость клеток Mono Mac 6, инфицированных Chlamydia pneumoniae, , зависит от активности связывания NF-κB.Заразить. Иммун. 69 : 7039-7045.
- 84.↵
Ward, M. E. 1999. Механизмы заболевания, вызванного хламидиозом, с. 171-210. В Р. С. Стивенс (ред.), Хламидии: внутриклеточная биология, патогенез и иммунитет. ASM Press, Вашингтон, округ Колумбия.
- 85.↵
Wehrl, W., V. Brinkmann, P. R. Jungblut, T. F. Meyer и A. J. Szczepek. 2004. Изнутри – процессинг хламидийного аутотранспортера PmpD и его роль в бактериальной адгезии и активации человеческих клеток-хозяев.Мол. Microbiol. 51 : 319-334.
- 86.↵
Xu, Q. 2003. Инфекции, белки теплового шока и атеросклероз. Curr. Opin. Кардиол. 18 : 245-252.
- 87.↵
Йошида, Н., К. Оэда, Э. Ватанабэ, Т. Миками, Ю. Фукита, К. Нисимура, К. Комаи и К. Мацуда. 2001. Шаперонин превратил токсин насекомых. Природа 411 : 44.
- 88.↵
Юань, Ю., К. Линг, Ю. X. Чжан, Д. Д. Рокки и Р. П. Моррисон. 1992. Моноклональные антитела определяют родоспецифические, видоспецифические и перекрестно-реактивные эпитопы хламидийного 60-килодальтонного белка теплового шока (hsp60): специфическое иммунодетектирование и очистка хламидийного hsp60. Заразить. Иммун. 60 : 2288-2296.
- 89.↵
Yucesan, C., and S. Sriram. 2001. Инфекция центральной нервной системы Chlamydia pneumoniae.